Not Merely Tau Tombstones, Neurofibrillary Tangles Are Dynamic
Quick Links
Far from being inert intracellular pebbles, inclusions formed by misfolded tau are quite dynamic, according to a paper in the January 26 Acta Neuropathologica. Todd Golde and colleagues at University of Florida, Gainesville, reported seeing turnover of tau inclusions in cultured slices of brain taken from young mice. Inclusions formed within 96 hours of seeding with tau fibrils, or infection with adenoviruses expressing mutant tau. The tau wandered in and out of the inclusions, with a half-life of about one week. As the aggregates aged, however, turnover slowed and half-life tripled. Knowing that inclusions are dynamic, researchers might find new ways to clear them, the authors suggested.
- Tau’s half-life in inclusions is about one week, longer in older inclusions.
- Inclusions seeded by mutant or wild-type tau turned over at same rate.
- Brain-slice model could be useful for studying inclusion-busting drugs.
“This article elegantly shows rapid turnover of soluble tau, while increasingly fibrillar tau shows increasingly slower turnover,” Benjamin Wolozin, Boston University, wrote to Alzforum (full comment below).
Previously, first author Cara Croft had collaborated with Benoit Giasson and colleagues, also at UF, to create a cell-culture model that did not require addition of tau fibrils to seed tangles (Strang et al., 2018). They infected cells with a recombinant adeno-associated virus (rAAV) containing human tau with two mutations, P301L and S320F, that cause frontotemporal dementia and parkinsonism linked to chromosome 17 (Rosso et al., 2002; Hutton et al., 1998). The scientists then used the same rAAV construct to infect organotypic mouse brain slices (Feb 2019 news). After 28 days, tau inclusions formed within neurons.
Now, Croft has tracked how those inclusions behave over time. She infected the brain slices with rAAVs containing human wild-type or P301L/S320F tau tagged with a Dendra2 motif. This fluorescent peptide is photoswitchable, meaning it changes irreversibly from emitting green photons to red after brief exposure to short-wavelength light. After this conversion, newly made tau can be distinguished by its green fluorescence, allowing the researchers to track new and older tau in real time.
In the slices, P301L/S320F tau formed inclusions within 96 hours and as quickly as 24 hours. After 10 days, about 87 percent of cells had inclusions of fibrillar tau, which lit up when stained with the β-sheet dye thiazine red. The Dendra-2-tagged tau clumped similarly to the unadulterated tau, indicating that the fluorescent motif was neither impeding nor aiding aggregation.
To track tau dynamics over time, Croft and colleagues cultured the transfected brain slices for 10 days, then shone light onto the cells to convert all the green-fluorescent tau to red. Then they repeatedly imaged the cells over 21 days, measuring how much tau was old versus new. Neurons produced wild-type and mutant tau at about the same rate. Wild-type formed no inclusions and had a half-life of about 2.7 days.
Tau in inclusions lasted longer, but not by much. New tau replaced old, gradually transitioning red clumps to green without affecting the size of the inclusion (see image below). Inclusion tau had a half-life of about eight days. Golde was surprised by how quickly tau turned over.
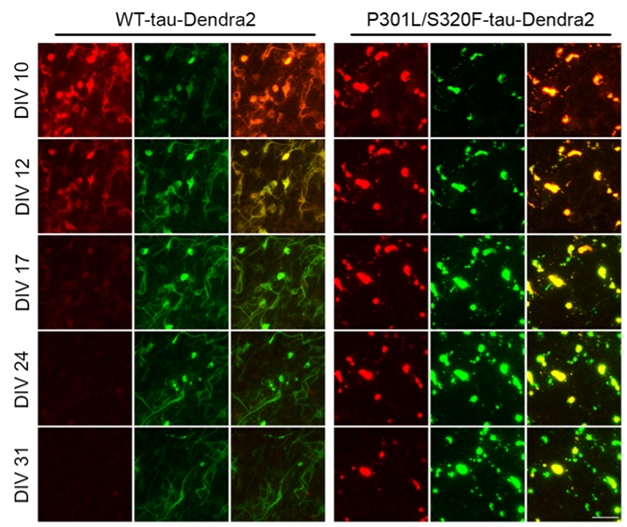
Tau Turnover. In mouse brain slices cultured for 10 days, green-fluorescent Dendra2-tau was zapped with light, turning the protein red (top row). Over the next 21 days, old tau turned over (left columns) as new tau formed (middle columns). Right columns show the mix. Mutant tau (right three columns) in inclusions lingered longer than did wild-type tau (left three columns) in the cytosol. [Courtesy of Croft et al., Acta Neuropathologica, 2021.]
Because tauopathies do not usually begin until midlife and inclusions are known to stick around for years, the authors wondered if tau dynamics changed as the tissue aged. The researchers cultured the brain slices for 30 or 60 days before converting the green-labeled tau to red, then measured turnover in the inclusions, again for at least 21 more days. The older the tissue culture, the longer tau’s half-life. It grew from eight days when photoswitching was done in 10-day-old slices to 16 days and 23 days when 30- and 60-day-old tissue was used, respectively. Even wild-type tau languished slightly longer as it aged, with a half-life of about four days in both 30- and 60-day-old slices, up from the 2.7 in 10-day-old tissue.
What about inclusions caused by fibrils of wild-type tau, such as those found in Alzheimer’s disease? Croft took fibrils made in Escherichia coli expressing the microtubule-binding domain of four-repeat (4R) tau and added them to 14-day-old slice cultures. These did not seed inclusions in wild-type tissue, but did seed in slices expressing P301L tau. P301L slices on their own did not form inclusions.
Inclusions formed within 96 hours of seeding. After 10 days, Croft shone light on the slices and measured inclusion turnover for another 10 days. Tau half-life was about eight days, or just the same as the P301L/S320F tau. In 30- and 60-day-old slices post-seeding, it was 18 and 24 days, respectively, again similar to half-lives cause by the double mutant.
Why tau turnover slows over time remains a mystery. Croft thinks that microglia and astrocytes may change over time, impeding tau clearance. Golde agreed, noting that tau turnover must be facilitated. “An isolated tangle does not spontaneously dissolve, so I do not see how it could be a passive process,” he told Alzforum.
Tara Spires-Jones, University of Edinburgh, was intrigued by the longer half-life in older cultures. “It begs the question of what the turnover is like in the aged human brain,” she wrote to Alzforum (full comment below). “Is this difference a function of the tissue or of the age of the inclusion?” Golde wondered the same thing. “There is evidence that cross-linking and other post-translational modifications occur in tau inclusions,” he said. “We can imagine that something cross-linked would be harder to remove.”
Marc Diamond, University of Texas Southwestern, Dallas, was not surprised by these findings. “We have known for a few decades that inclusions are not ‘rocks’ but can be disassembled,” he wrote to Alzforum. “Presumably the aggregates get more consolidated, as other proteins pile on, and it is a bigger mess to disassemble.”
Previously, researchers in David Holtzman’s lab at Washington University School of Medicine, St. Louis, examined tau turnover in the brains of transgenic mice (Yamada et al., 2015). Soluble tau turned over faster than insoluble tau, with half-lives of about 10 and 34 days, respectively. Phosphorylated soluble tau cleared even faster, with half-lives between five to 10 days, depending on which amino acid was modified. The authors noted that insoluble tau likely represented oligomers, fibrils, and other aggregates, but they did not look at inclusions specifically.
“Overall, our findings are reasonably consistent with Croft’s despite using very different methods,” Holtzman told Alzforum. “We both show that tau aggregates are in equilibrium with soluble tau.”
Also at Wash U, Randy Bateman, Celeste Karch, and colleagues studied tau turnover in human induced pluripotent stem cell (iPSC)-derived neurons (March 2018 news). Using stable isotope labeling, the researchers tracked how much tau was present over time via mass spectrometry. In cultured neurons, tau’s half-life was seven days, with phosphorylated and 4R tau turning over quicker than the three-repeat variety.
What about other tau mutations or other tauopathies besides FTD and PD? Michel Goedert, MRC Laboratory of Molecular Biology, Cambridge, England, U.K., noted that inclusion turnover rates likely depend on filament structures and modifications of their components, which differ between diseases. “For example, tau filament folds differ among AD, chronic traumatic encephalopathy, FTDP-17, and corticobasal degeneration,” he wrote to Alzforum (full comment below).
Holtzman proposed that because of this ebb and flow of tau, one could possibly get rid of the aggregates by removing the surrounding soluble tau. Claire Durrant, University of Edinburgh, warned against potentially unwanted effects of this, such as releasing toxic, synapse-disrupting oligomers. “This model could be useful for testing this principle, by correlating increased or decreased turnover with measures of neuronal health and synaptic function,” she wrote to Alzforum (full comment below). Diamond agreed. “This is a strong system to study the biological basis of aggregate turnover, and could have important implications for identifying factors involved in this process.”—Chelsea Weidman Burke
References
News Citations
- Viral Vectors Trigger Robust Tauopathy in Brain Slices
- Isotope Labeling Links Tau Production to Aβ Burden
Paper Citations
- Strang KH, Croft CL, Sorrentino ZA, Chakrabarty P, Golde TE, Giasson BI. Distinct differences in prion-like seeding and aggregation between Tau protein variants provide mechanistic insights into tauopathies. J Biol Chem. 2018 Feb 16;293(7):2408-2421. Epub 2017 Dec 19 PubMed.
- Rosso SM, van Herpen E, Deelen W, Kamphorst W, Severijnen LA, Willemsen R, Ravid R, Niermeijer MF, Dooijes D, Smith MJ, Goedert M, Heutink P, van Swieten JC. A novel tau mutation, S320F, causes a tauopathy with inclusions similar to those in Pick's disease. Ann Neurol. 2002 Mar;51(3):373-6. PubMed.
- Hutton M, Lendon CL, Rizzu P, Baker M, Froelich S, Houlden H, Pickering-Brown S, Chakraverty S, Isaacs A, Grover A, Hackett J, Adamson J, Lincoln S, Dickson D, Davies P, Petersen RC, Stevens M, de Graaff E, Wauters E, van Baren J, Hillebrand M, Joosse M, Kwon JM, Nowotny P, Che LK, Norton J, Morris JC, Reed LA, Trojanowski J, Basun H, Lannfelt L, Neystat M, Fahn S, Dark F, Tannenberg T, Dodd PR, Hayward N, Kwok JB, Schofield PR, Andreadis A, Snowden J, Craufurd D, Neary D, Owen F, Oostra BA, Hardy J, Goate A, van Swieten J, Mann D, Lynch T, Heutink P. Association of missense and 5'-splice-site mutations in tau with the inherited dementia FTDP-17. Nature. 1998 Jun 18;393(6686):702-5. PubMed.
- Yamada K, Patel TK, Hochgräfe K, Mahan TE, Jiang H, Stewart FR, Mandelkow EM, Holtzman DM. Analysis of in vivo turnover of tau in a mouse model of tauopathy. Mol Neurodegener. 2015 Oct 26;10:55. PubMed.
Further Reading
Primary Papers
- Croft CL, Goodwin MS, Ryu DH, Lessard CB, Tejeda G, Marrero M, Vause AR, Paterno G, Cruz PE, Lewis J, Giasson BI, Golde TE. Photodynamic studies reveal rapid formation and appreciable turnover of tau inclusions. Acta Neuropathol. 2021 Mar;141(3):359-381. Epub 2021 Jan 26 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Edinburgh
This paper by Dr. Cara Croft and colleagues is an elegant demonstration that fibrillar tau aggregates in neurons are not static structures in the organotypic brain slice model used. I was particularly interested that the half-life of inclusions was longer in longer-term cultures, which begs the questions, what is the turnover like in aged human brain? Is this difference a function of the age of the tissue or age of the inclusion?
This paper is important as it lends hope that strategies to prevent tau aggregation may both prevent further aggregation and eventually lead to removal of existing aggregates due to this turnover. However, it is worth a note of caution that it is likely not the fibrillar aggregates that are toxic but rather soluble forms of tau, so we need a clearer understanding of which forms of tau are neurotoxic in order to develop effective therapeutic strategies.
MRC Laboratory of Molecular Biology
Abundant filamentous tau inclusions are characteristic of a number of human neurodegenerative diseases, collectively referred to as tau proteinopathies. The presence of inclusions correlates with neurodegeneration and a causal connection became clear following the identification of mutations in MAPT that give rise to familial forms of frontotemporal dementia.
Croft and colleagues showed previously that brain-slice cultures transduced with recombinant adeno-associated viruses that encode 0N4R P301L/S320F human tau develop inclusions and subsequent nerve cell death (Croft et al., 2019). Abundant tau filaments were present. In the new work, they now report that the inclusions are dynamic, with a half-life of approximately seven days (Croft et al., 2021).
The turnover rates of protein inclusions probably depend on the structures of the constituent filaments, their modifications and association with other molecules. In Alzheimer’s disease (AD) and chronic traumatic encephalopathy (CTE), extracellular tangles, so-called “tombstones,” predominate at end-stage. This is not true of Pick’s disease (PiD) or corticobasal degeneration (CBD). Over the past four years, we used electron cryo-microscopy to show that tau filament folds differ between AD, CTE, PiD and CBD, with those of AD and CTE being most similar (Fitzpatrick et al., 2017; Falcon et al., 2018; Falcon et al., 2019; Zhang et al., 2020). Despite these structural differences, tau filaments from brain comprise at least microtubule-binding repeats 3 (R3) and R4, as well as 10-12 amino acids after R4. Constructs K18 and K19 of recombinant tau end at E372, four amino acids after R4 (Gustke et al., 1994). Even though the structures of K18 and K19 filaments are not known, they probably differ from those of tau filaments from human brain.
The structures of tau filaments from cases with mutations in MAPT are also unknown. As in CBD, 4R tau assembles in some nerve cells and glial cells in individuals with the P301L mutation, with only a small number of extracellular inclusions (Spillantini et al., 1998). Less is known about the effects of mutation S320F (Rosso et al., 2002). There are no known cases of frontotemporal dementia that are caused by both missense mutations. Even so, it may be interesting to know more about the structures and modifications of 0N4R P301L/S320F human tau filaments.
References:
Croft CL, Cruz PE, Ryu DH, Ceballos-Diaz C, Strang KH, Woody BM, Lin WL, Deture M, Rodríguez-Lebrón E, Dickson DW, Chakrabarty P, Levites Y, Giasson BI, Golde TE. rAAV-based brain slice culture models of Alzheimer's and Parkinson's disease inclusion pathologies. J Exp Med. 2019 Mar 4;216(3):539-555. Epub 2019 Feb 15 PubMed.
Falcon B, Zhang W, Murzin AG, Murshudov G, Garringer HJ, Vidal R, Crowther RA, Ghetti B, Scheres SH, Goedert M. Structures of filaments from Pick's disease reveal a novel tau protein fold. Nature. 2018 Sep;561(7721):137-140. Epub 2018 Aug 29 PubMed.
Falcon B, Zivanov J, Zhang W, Murzin AG, Garringer HJ, Vidal R, Crowther RA, Newell KL, Ghetti B, Goedert M, Scheres SH. Novel tau filament fold in chronic traumatic encephalopathy encloses hydrophobic molecules. Nature. 2019 Apr;568(7752):420-423. Epub 2019 Mar 20 PubMed.
Fitzpatrick AW, Falcon B, He S, Murzin AG, Murshudov G, Garringer HJ, Crowther RA, Ghetti B, Goedert M, Scheres SH. Cryo-EM structures of tau filaments from Alzheimer's disease. Nature. 2017 Jul 13;547(7662):185-190. Epub 2017 Jul 5 PubMed.
Gustke N, Trinczek B, Biernat J, Mandelkow EM, Mandelkow E. Domains of tau protein and interactions with microtubules. Biochemistry. 1994 Aug 16;33(32):9511-22. PubMed.
Rosso SM, van Herpen E, Deelen W, Kamphorst W, Severijnen LA, Willemsen R, Ravid R, Niermeijer MF, Dooijes D, Smith MJ, Goedert M, Heutink P, van Swieten JC. A novel tau mutation, S320F, causes a tauopathy with inclusions similar to those in Pick's disease. Ann Neurol. 2002 Mar;51(3):373-6. PubMed.
Spillantini MG, Crowther RA, Kamphorst W, Heutink P, van Swieten JC. Tau pathology in two Dutch families with mutations in the microtubule-binding region of tau. Am J Pathol. 1998 Nov;153(5):1359-63. PubMed.
Zhang W, Tarutani A, Newell KL, Murzin AG, Matsubara T, Falcon B, Vidal R, Garringer HJ, Shi Y, Ikeuchi T, Murayama S, Ghetti B, Hasegawa M, Goedert M, Scheres SH. Novel tau filament fold in corticobasal degeneration. Nature. 2020 Apr;580(7802):283-287. Epub 2020 Feb 12 PubMed.
This is a really interesting paper and provides a potentially very useful new tool for studying tau inclusions. Whilst tau tangles have long been reported as a key feature in Alzheimer's disease and other tauopathies, we know surprisingly little about them! As the authors discuss in the paper, we know that tangles can exist for long periods of time within neurons, but the impact on those neurons is somewhat unclear, despite tau tangles being a reasonable correlate of cognitive symptoms in disease. There have been previous reports (e.g. Dejanovic et al., 2018) that show that high levels of phosphorylated tau can result in synapses being labelled by the complement system—targeting them for destruction by microglia—potentially explaining the reduction in synapse density in tauopathies. Other papers, however (Kuchibhotla et al., 2014; Rocher et al., 2010; Rudinskiy et al., 2014), find that tangle-bearing neurons are relatively functional, and can even integrate into functional neuronal circuits. This could indicate that other tau forms, such as soluble oligomers, could be more toxic to the cell.
The authors use brain-slice cultures, which are an effective ex vivo tool that permits ease of manipulation (such as transfection with viruses and live imaging), in a complex environment that, at least partially, retains neuronal circuits, synaptic connections, and supporting cell types (such as microglia, astrocytes, etc.). It is worth noting that whilst the tissue used is postnatal, this group (Croft et al., 2017, and others, myself included, e.g., Harwell and Coleman, 2016; Durrant et al., 2020) find that AD-like pathological phenotypes partially replicate and occur on an accelerated timescale in slice cultures relative to in vivo.
This paper addresses a key question about the nature of tau tangles—reporting that these structures are far more dynamic than originally thought. The methodology is neat, using a photo-convertible tau Dendra2 virus to follow turnover of tau as well as inclusion of new protein in the same tangles over time. Their finding that tau tangles are dynamic structures could highlight the potential for increasing tau clearance from the brain and reducing tangle burden if the mechanisms behind turnover can be uncovered.
One point worth considering, however, is whether, given the assertion in this and other studies that tau tangles themselves may not be overtly harmful to the neurons, clearance of tangles could result in unwanted side effects, such as release of toxic oligomers that are known to have synapse-disrupting properties. This model could provide a useful mechanism for testing this principle, by correlating increased or decreased turnover with measures of neuronal health/synaptic function. The platform would also be a relatively quick and effective drug screening tool for compounds looking to disrupt tau aggregation, or promote tau clearance.
Interestingly, only mutant tau is found to form aggregates in the culture period, likely reinforcing the dogma that hyperphosphorylated tau is the main component of such tangle structures. Whilst the mutant tau is a very useful experimental tool for this system, it is worth noting that most individuals with tau tangles do not possess tau mutations—so whilst mechanisms are very likely to be similar, extrapolation to the behavior of tangles composed of "wild-type" tau may require additional experimentation.
The authors maintained the cultures for an impressive length of time (up to 100 days in some cases) and found that the half-life of tau inclusions increased with the increasing age of the slice culture. It would be very interesting to uncover whether this is a sign of reduced slice health due to culture conditions, or a genuine marker of neuronal age showing reduced capacity for tau turnover. If the latter, this could provide a very interesting direction to explore whether age alters responses to tau pathology, or even predisposes to formation of tangles in the first place due to reduced clearance of the aggregates. This could open up understanding of why humans are more susceptible to tauopathies with increasing age.
Overall, I think the authors present an excellent new tool for exploring tau-tangle turnover mechanisms in neurons and have provided novel insight into the dynamic nature of tau inclusions.
References:
Dejanovic B, Huntley MA, De Mazière A, Meilandt WJ, Wu T, Srinivasan K, Jiang Z, Gandham V, Friedman BA, Ngu H, Foreman O, Carano RA, Chih B, Klumperman J, Bakalarski C, Hanson JE, Sheng M. Changes in the Synaptic Proteome in Tauopathy and Rescue of Tau-Induced Synapse Loss by C1q Antibodies. Neuron. 2018 Dec 19;100(6):1322-1336.e7. Epub 2018 Nov 1 PubMed.
Kuchibhotla KV, Wegmann S, Kopeikina KJ, Hawkes J, Rudinskiy N, Andermann ML, Spires-Jones TL, Bacskai BJ, Hyman BT. Neurofibrillary tangle-bearing neurons are functionally integrated in cortical circuits in vivo. Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):510-4. Epub 2013 Dec 24 PubMed.
Rocher AB, Crimins JL, Amatrudo JM, Kinson MS, Todd-Brown MA, Lewis J, Luebke JI. Structural and functional changes in tau mutant mice neurons are not linked to the presence of NFTs. Exp Neurol. 2010 Jun;223(2):385-93. PubMed.
Rudinskiy N, Hawkes JM, Wegmann S, Kuchibhotla KV, Muzikansky A, Betensky RA, Spires-Jones TL, Hyman BT. Tau pathology does not affect experience-driven single-neuron and network-wide Arc/Arg3.1 responses. Acta Neuropathol Commun. 2014 Jun 10;2:63. PubMed.
Croft CL, Wade MA, Kurbatskaya K, Mastrandreas P, Hughes MM, Phillips EC, Pooler AM, Perkinton MS, Hanger DP, Noble W. Membrane association and release of wild-type and pathological tau from organotypic brain slice cultures. Cell Death Dis. 2017 Mar 16;8(3):e2671. PubMed.
Harwell CS, Coleman MP. Synaptophysin depletion and intraneuronal Aβ in organotypic hippocampal slice cultures from huAPP transgenic mice. Mol Neurodegener. 2016 Jun 10;11(1):44. PubMed.
Durrant CS, Ruscher K, Sheppard O, Coleman MP, Özen I. Beta secretase 1-dependent amyloid precursor protein processing promotes excessive vascular sprouting through NOTCH3 signalling. Cell Death Dis. 2020 Feb 6;11(2):98. PubMed.
Make a Comment
To make a comment you must login or register.