Amyloid Jungle: Plaque Fibrils Mesh With All Manner of Vesicles, Membranes
Quick Links
Scientists have long known that amyloid plaques consist of more than just Aβ. With cryo-electron tomography, they can now zoom in on these aggregated jumbles, revealing in startling detail the mess they truly are. On May 17 in Nature Communications, Sheena Radford, Neil Ranson, and René Frank at the University of Leeds, U.K., and colleagues describe their “in tissue” study. It spied Aβ fibrils arranged in parallel bundles and lattices in APPNL-G-F knock-in mice. It also spotted thinner filaments, and highly branched fibrils, within the plaque. The big surprise? Interwoven with these different forms were remnants of subcellular compartments, such as extracellular vesicles, droplets, and multilamellar bodies. Cryo-EM showed that individual Aβ fibrils were composed of two S-shaped protofilaments. Although similar to previously reported structures of Aβ42 isolated from human brain, the Arctic mutation introduced substantial kinks.
- Cryo-electron tomography charts molecular structure of plaques in situ.
- Aβ42 fibrils of varying sizes mingle with vesicular components.
- Extracellular vesicles of different shapes and sizes abound.
- Cryo-EM unveiled a paired, S-shaped protofilament core of Aβ42 fibrils containing the Arctic mutation.
“It reminds us that the amyloid plaque is not simply an inert ball of fibrils—it is a dynamic structure with many pathologic cellular events happening within and nearby,” wrote Dennis Selkoe and Andrew Stern of Brigham and Women’s Hospital in Boston. The variety of amyloid fibrils and the many membranous compartments spotted within plaques underscore the malignancy of Aβ deposition and emphasize the importance of preventing this “multicellular debacle” in the brain, they added (comment below).
“The work … represents a veritable tour de force,” wrote Michel Goedert and Sjors Scheres of MRC Laboratory of Molecular Biology, Cambridge, U.K. (comment below).
Advances in cryo-EM have graced the field with structures of amyloid folds in recent years, including the S-shaped protofilaments that buddy up to form the core of Aβ42 fibrils in the AD brain (Jan 2022 news). While atomic structures dazzle with high resolution, they provide no spatial context for how these fibrils sit within the brain parenchyma, or whose company they keep inside a plaque.
To examine the molecular architecture of Aβ plaques in APPNL-G-F mice, first author Conny Leistner and colleagues used cryo-correlated light and electron microscopy (cryo-CLEM) to guide them toward fluorescently labeled Aβ plaques within frozen brain sections, then applied cryo-electron tomography.
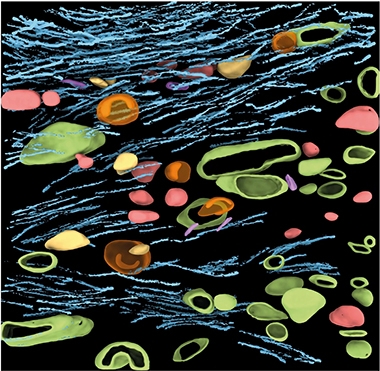
Innards of a Plaque. Bundles of Aβ fibrils (blue) mingle with round (red), C-shaped (orange), and ellipsoid (purple) vesicles, as well as membrane-less droplets (yellow). Plasma membrane from intact neighboring cells, or from cellular remnants (green) interdigitate. [Courtesy of Leistner et al., Nature Communications, 2023.]
Across the 23 tomograms, or maps, of Aβ plaques that the researchers collected, they spotted dense arrays of fibrils, arranged as lattices, or in parallel bundles. Within plaque cores, these fibrils were interspersed with vesicles of three varieties: round, ellipsoid, and EVs carrying cup or C-shaped membranes within their lumens—in other words, vesicles within vesicles. Multilamellar bodies, which are vesicles wrapped in multiple concentric rings of membrane, showed up in nearly a third of tomograms. These multimembrane-encased vesicles are thought to be produced during autophagy, and have been spotted among dystrophic neurites in the AD brain (Hariri et al., 2000; Nixon et al., 2005). Small droplets also appeared within the plaques. At 80-120 nm in size, the smooth, spherical structures resembled lipid droplets known to reside within healthy cells, but were five to 10 times smaller (Klumpe et al., 2021). The researchers confirmed that these plaques and their subcellular contents were entirely extracellular, although plasma membranes of intact neighboring cells contacted the plaque surface. They also spotted plasma membranes within the core of the plaques, which Frank said could derive from processes of intact cells poking deep into the plaques, and/or from bits of membrane broken off from perished cells (see image above).
Where did all these membranous bubbles come from, and how did they get mixed up with Aβ plaques? This question remains, Frank told Alzforum. Tsuneya Ikezu of the Mayo Clinic in Jacksonville, Florida, thinks the vesicles may have come from surrounding activated glia. “Indeed, the ultrastructural analysis of the subcellular and membrane-bound compartments surrounding the MX04-stained plaques are consistent with those of microglia and astrocytes,” he wrote” (comment below).
Ikezu's group identified plaque-adjacent microglia churning out EVs in the same mouse model (Clayton et al., 2021). Ikezu thinks the multilamellar bodies within some plaques could derive from intermediates of autophagy in the surrounding glia, noting that similar multilayer vesicles have been seen in human cerebrospinal fluid (Emelyanov et al., 2020).
Could some vesicles come from neurons? Ralph Nixon of New York University told Alzforum that the components are consistent with PANTHOS, aka perinuclear rosettes of Aβ fibrils mixed with ER, Golgi, and lysosomal membranes, which show up in some neurons prior to plaque formation (Jun 2022 news). Specific labeling of the proteins involved would be needed to be sure. PANTHOS-derived Aβ plaques form when their host neurons burst. Frank noted that such neurodegeneration is not a feature of the APPNL-G-F mouse model.
“To figure out the source of non-amyloid constituents [in the plaques], we think it is important to obtain tomograms at earlier stages of β-amyloid biogenesis,” Frank told Alzforum. Gunnar Gouras of Lund University in Sweden, whose work suggests that plaque formation begins inside cells, noted that at these early stages, the researchers may need to hunt for structures containing Aβ oligomers, which may not be labeled with amyloid dyes.
Focus on Fibrils
Besides viewing the molecular architecture of plaques with cryo-ET, Leistner and colleagues used cryo-EM to resolve the atomic details of individual fibrils purified from plaques. At a resolution of 3.0 angstroms, a predominant, left-handed twist emerged. The ordered core of these fibrils comprised a pair of S-shaped protofilaments, tethered by an extensive interface (image below). Residues 1-38 from each Aβ42 peptide contributed to this S-shaped core. Though similar to the type I and II amyloid folds found previously in sporadic AD, and familial AD brain samples, respectively, these APPNL-G-F mouse fibrils were a bit different, likely owing to the presence of the E22G Arctic mutation. The amino acid change allowed residue G22 to become deeply buried within the fibril structure, compared with its exposed position in wild-type fibrils (Yang et al., 2023).

Another S. The protofilament core of an Aβ fibril from APPNL-G-F mouse featured two S-shaped protofilaments nestled together. Residue G22, the Arctic mutation, is buried within the fibril structure. [Courtesy of Leistner et al., Nature Communications, 2023.]
Scheres and Goedert noted that the structure is a spot-on match with what they observed via cryo-EM in older APPNL-G-F mice (Yang et al., 2023). They also confirmed that, owing to the presence of the Arctic mutation, these fibrils differed from those prepared from APPNL-F mice, which, in turn, matched with Type II human filaments (Yang et al., 2023).
While only one predominant Aβ42 fibril structure could be resolved at atomic resolution, the researchers noticed several forms of amyloid in their tomograms. These included thinner filaments, of about half the width of those observed via cryo-EM, which the scientists called “protofilament-like rods.” They tended to branch from thicker parent fibrils, sometimes forming a bridge between them. By tracing individual filaments within parallel bundles in the tomograms, the researchers saw branched fibrils intermingled with unbranched, the latter appearing to match the structure resolved via cryo-EM (image below). These various forms did not match the predicted worm-like protofibrils thought to be the target of lecanemab—a therapeutic antibody raised against Aβ42 with the Arctic mutation (Aug 2001 news).

Variety Pack. A bundle of parallel fibrils traced from a tomogram. Magenta arrows indicate putative fibril branch points; blue arrows point to unbranched fibrils. [Courtesy of Leistner et al., Nature Communications, 2023.]
Selkoe and Stern questioned the identity of these other amyloids. “Without further evidence, such as immunolabeling or higher-resolution structure determination, we cannot yet conclude that these thinner or branched structures, which are a minority among the abundant amyloid fibrils, are composed of Aβ,” they wrote.
In the same vein, Colin Masters of the University of Melbourne noted that the Methoxy-X04 dye the authors used to label amyloid is known to bind any structure containing β-pleated sheets. “Uncertainties regarding the observed fibrillar/filamentous aggregates could be addressed through immunolabeling EM, particularly by employing lecanemab,” he noted. “It remains unclear whether the observed ‘in-tissue’ molecular architecture includes exclusively Aβ fibrillar forms or if other β-sheet-rich species (such as tau) or globular (co)-aggregates contribute to the lesions of AD,” he wrote (comment below).—Jessica Shugart
References
News Citations
- Cryo-EM Unveils Distinct Aβ42 Fibril Structures for Sporadic, Familial AD
- Behold PANTHOS, a Toxic Wreath of Perinuclear Aβ That Kills Neurons
- "Arctic" APP Mutation Supports Protofibril Role
Research Models Citations
Mutations Citations
Paper Citations
- Hariri M, Millane G, Guimond MP, Guay G, Dennis JW, Nabi IR. Biogenesis of multilamellar bodies via autophagy. Mol Biol Cell. 2000 Jan;11(1):255-68. PubMed.
- Nixon RA, Wegiel J, Kumar A, Yu WH, Peterhoff C, Cataldo A, Cuervo AM. Extensive involvement of autophagy in Alzheimer disease: an immuno-electron microscopy study. J Neuropathol Exp Neurol. 2005 Feb;64(2):113-22. PubMed.
- Klumpe S, Fung HK, Goetz SK, Zagoriy I, Hampoelz B, Zhang X, Erdmann PS, Baumbach J, Müller CW, Beck M, Plitzko JM, Mahamid J. A modular platform for automated cryo-FIB workflows. Elife. 2021 Dec 24;10 PubMed.
- Clayton K, Delpech JC, Herron S, Iwahara N, Ericsson M, Saito T, Saido TC, Ikezu S, Ikezu T. Plaque associated microglia hyper-secrete extracellular vesicles and accelerate tau propagation in a humanized APP mouse model. Mol Neurodegener. 2021 Mar 22;16(1):18. PubMed. Correction.
- Emelyanov A, Shtam T, Kamyshinsky R, Garaeva L, Verlov N, Miliukhina I, Kudrevatykh A, Gavrilov G, Zabrodskaya Y, Pchelina S, Konevega A. Cryo-electron microscopy of extracellular vesicles from cerebrospinal fluid. PLoS One. 2020;15(1):e0227949. Epub 2020 Jan 30 PubMed.
- Yang Y, Arseni D, Zhang W, Huang M, Lövestam S, Schweighauser M, Kotecha A, Murzin AG, Peak-Chew SY, Macdonald J, Lavenir I, Garringer HJ, Gelpi E, Newell KL, Kovacs GG, Vidal R, Ghetti B, Ryskeldi-Falcon B, Scheres SH, Goedert M. Cryo-EM structures of amyloid-β 42 filaments from human brains. Science. 2022 Jan 14;375(6577):167-172. Epub 2022 Jan 13 PubMed.
- Yang Y, Zhang W, Murzin AG, Schweighauser M, Huang M, Lövestam S, Peak-Chew SY, Saito T, Saido TC, Macdonald J, Lavenir I, Ghetti B, Graff C, Kumar A, Nordberg A, Goedert M, Scheres SH. Cryo-EM structures of amyloid-β filaments with the Arctic mutation (E22G) from human and mouse brains. Acta Neuropathol. 2023 Mar;145(3):325-333. Epub 2023 Jan 7 PubMed.
Further Reading
Primary Papers
- Leistner C, Wilkinson M, Burgess A, Lovatt M, Goodbody S, Xu Y, Deuchars S, Radford SE, Ranson NA, Frank RA. The in-tissue molecular architecture of β-amyloid pathology in the mammalian brain. Nat Commun. 2023 May 17;14(1):2833. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
Brigham and Women's Hospital, Harvard Medical School
Co-Director, Brigham and Women's Hospital's Ann Romney Center for Neurologic Diseases
This is a beautiful ultrastructural study from the Frank lab. As early as the 1960s, electron microscopy of Alzheimer’s disease brains (and later brains of mouse models) revealed dystrophic neurites, multilaminar vesicles, autophagic vacuoles, and other abnormal cellular structures intertwined with extracellular amyloid fibrils (e.g., Kidd, 1964; Yamaguchi et al., 1989; Masliah et al., 1996). However, these studies relied on heavy fixation and two-dimensional analysis. Now, Leistner et al. use cryo-electron tomography correlated with fluorescence microscopy to generate elegant three-dimensional maps of amyloid plaques in fresh Arctic mutant APP knock-in mouse brain tissue without fixation. The rich array of structures present in close apposition to Aβ fibrils (e.g. Figs 2 and 3) is impressive. It reminds us that the amyloid plaque is not simply an inert ball of fibrils—it is a dynamic structure with many pathologic cellular events happening within and nearby.
The authors make special efforts to show that the amyloid fibrils are extracellular, not within the many subcellular organelles in the intimately associated cytoplasm, clarifying a recurring question about whether intracellular loci are common sites of Aβ accrual. Further, they show that isolated ex vivo preparations of fibrils (e.g., in sarkosyl extracts) are not identical to those they painstakingly visualize in fresh, intact brain.
We disagree, at this juncture, with the conclusions reached from Figure 6, regarding the presence of branched, thin protofilaments. We reserve the term “protofilament” to have a strict definition in the context of AD: a single stack of misfolded Aβ in a β-pleated sheet conformation. In the case of Aβ, two protofilaments form one filament (or fibril). The 3-5 nm branched structures that Leistner et al. observe could be composed of Aβ, but they might instead be composed of any elongated macromolecules interspersed with bona fide Aβ fibrils: other proteins, carbohydrates, nucleic acids, or some complex of these.
For instance, amyloid plaques have been shown to stain intensely for RNA (Ginsberg et al.,1997). Without further evidence, such as immunolabeling or higher-resolution structure determination, we cannot yet conclude that these thinner or branched structures, which are a minority among the abundant amyloid fibrils, are composed of Aβ. In aqueous extracts prepared by simple diffusion (without homogenization or sarkosyl) from typical AD brains, we have not observed any elongated 3-5 nm structures labeling with Aβ antibodies, but it is certainly possible these were lost during preparation (Stern et al., 2023). We look forward to further advances in cryoET and related techniques giving us clearer pictures of diverse Aβ fibrils and associated structures.
The complexity and heterogeneity of the amyloid fibrils that accumulate in AD brain, and the many altered intracellular organelles and extracellular membranous structures documented by Leistner et al. in their cryoET reconstructions, underscore the malignancy of Aβ deposition and emphasize the importance of preventing its deposition in the first place. Prevention will likely be far more successful than attempting to arrest and repair this multicellular debacle in the brain.
References:
Kidd M. ALZHEIMER'S DISEASE--AN ELECTRON MICROSCOPICAL STUDY. Brain. 1964 Jun;87:307-20. PubMed.
Yamaguchi H, Nakazato Y, Hirai S, Shoji M, Harigaya Y. Electron micrograph of diffuse plaques. Initial stage of senile plaque formation in the Alzheimer brain. Am J Pathol. 1989 Oct;135(4):593-7. PubMed.
Masliah E, Sisk A, Mallory M, Mucke L, Schenk D, Games D. Comparison of neurodegenerative pathology in transgenic mice overexpressing V717F beta-amyloid precursor protein and Alzheimer's disease. J Neurosci. 1996 Sep 15;16(18):5795-811. PubMed.
Ginsberg SD, Crino PB, Lee VM, Eberwine JH, Trojanowski JQ. Sequestration of RNA in Alzheimer's disease neurofibrillary tangles and senile plaques. Ann Neurol. 1997 Feb;41(2):200-9. PubMed.
Stern AM, Yang Y, Jin S, Yamashita K, Meunier AL, Liu W, Cai Y, Ericsson M, Liu L, Goedert M, Scheres SH, Selkoe DJ. Abundant Aβ fibrils in ultracentrifugal supernatants of aqueous extracts from Alzheimer's disease brains. Neuron. 2023 Jul 5;111(13):2012-2020.e4. Epub 2023 May 10 PubMed.
MRC Laboratory of Molecular Biology
MRC Laboratory of Molecular Biology
This work represents a veritable tour de force. The authors used high-pressure freezing to preserve fresh brains from AppNL-G-F knock-in mice for fluorescence light microscopy and electron cryo-tomography (cryo-ET). These mice deposit abundant Aβ42 plaques that are made of human Aβ with the Arctic mutation E22G (Saito et al., 2014). Leistner et al. used the fluorescence of Aβ plaques labelled by the amyloid dye methoxy-X04 to know where to cut the frozen brain tissues into sections thin enough for transmission electron microscopy, and then performed cryo-ET on those sections.
This work is pioneering the characterization of rodent brains with human disease pathologies at the structural molecular level. It describes the three-dimensional arrangement of Aβ filaments in situ. Ultimately, one would want to be able to perform this type of analysis in human brains from individuals with various neurodegenerative diseases. Knowledge of where amyloid filaments are in the brain and how they interact with other cellular components will be invaluable, not only for understanding fundamental aspects of human diseases, but also for examining the validity of model systems.
The authors also extracted Aβ42 filaments from the brains of 11- to 13-month-old homozygous AppNL-G-F mice and performed standard helical reconstruction electron cryo-microscopy (cryo-EM) to obtain an atomic model. Filament structures were identical to those we determined previously from the brains of 22-month-old homozygous AppNL-G-F mice, demonstrating that the filament structures did not change between the ages of 11 and 22 months (Yang et al., 2023).
We also reported that the structures of Aβ42 filaments from the temporal cortex of an individual with the Arctic mutation differed from those of AppNL-G-F knock-in mice, raising doubts about the usefulness of this line. Homozygous mice from knock-in line AppNL-F deposit wild-type human Aβ42 (Saito et al., 2014) that is identical in structure to Type II Aβ42 filaments from the brains of individuals with Alzheimer’s disease or with a variety of other diseases with Aβ42 co-pathology (Yang et al., 2022). It will be interesting to study the molecular architecture of Aβ path biology in the brains from AppNL-F mice.
References:
Saito T, Matsuba Y, Mihira N, Takano J, Nilsson P, Itohara S, Iwata N, Saido TC. Single App knock-in mouse models of Alzheimer's disease. Nat Neurosci. 2014 May;17(5):661-3. Epub 2014 Apr 13 PubMed.
Yang Y, Arseni D, Zhang W, Huang M, Lövestam S, Schweighauser M, Kotecha A, Murzin AG, Peak-Chew SY, Macdonald J, Lavenir I, Garringer HJ, Gelpi E, Newell KL, Kovacs GG, Vidal R, Ghetti B, Ryskeldi-Falcon B, Scheres SH, Goedert M. Cryo-EM structures of amyloid-β 42 filaments from human brains. Science. 2022 Jan 14;375(6577):167-172. Epub 2022 Jan 13 PubMed.
Yang Y, Zhang W, Murzin AG, Schweighauser M, Huang M, Lövestam S, Peak-Chew SY, Saito T, Saido TC, Macdonald J, Lavenir I, Ghetti B, Graff C, Kumar A, Nordberg A, Goedert M, Scheres SH. Cryo-EM structures of amyloid-β filaments with the Arctic mutation (E22G) from human and mouse brains. Acta Neuropathol. 2023 Mar;145(3):325-333. Epub 2023 Jan 7 PubMed.
Mayo Clinic Florida
I find this study quite exciting to visualize the amyloid fibrils in the brains of APPNL-G-F knock-in mice using cryo-electron tomography (Cryo-ET). One of the striking findings is the identification of so many extracellular vesicles/exosomes (50-200 nm in diameter) in the plaque region, which are 100-fold more abundant than in wild-type mouse brain. This study demonstrates that EVs accumulate in the amyloid plaques. The EVs could originate from surrounding activated glia. Indeed, the ultrastructural analysis of the sub-cellular and membrane-bound compartments surrounding the MX04-stained plaques is consistent with those of microglia and astrocytes. This is in line with our recent study, showing that plaque-associated microglia are activated and exhibit enhanced biogenesis and secretion of EVs, which we visualized in single-vesicle resolution in the APPNL-G-F knock-in mouse brain (Clayton et al., 2021).
In accord, one of the well-accepted EV markers, CD9, is also a marker of disease-associated / neurodegenerative microglia. The authors also identified multilamellar bodies composed of vesicles wrapped with multiple concentric rings of membrane, which they discussed as intracellular intermediates of autophagy. A similar structure was also identified in the EVs isolated from human cerebrospinal fluid by Cryo-EM (Emelyanov et al., 2020), suggesting that autophagy intermediate-derived EVs may also be secreted from the surrounding activated glia.
It would be of interest, if these findings can be reproduced in other APP mouse models or human AD brain tissues, to know how the depletion of microglia, or their inactivation, alters the EV deposition around plaques, and if these EVs also contain tau aggregates, as recently shown by the Duff and Ryskeldi-Falcon groups (Fowler et al., 2023). These EVs may also be the source of dystrophic neurites and other pathologies.
References:
Clayton K, Delpech JC, Herron S, Iwahara N, Ericsson M, Saito T, Saido TC, Ikezu S, Ikezu T. Plaque associated microglia hyper-secrete extracellular vesicles and accelerate tau propagation in a humanized APP mouse model. Mol Neurodegener. 2021 Mar 22;16(1):18. PubMed. Correction.
Emelyanov A, Shtam T, Kamyshinsky R, Garaeva L, Verlov N, Miliukhina I, Kudrevatykh A, Gavrilov G, Zabrodskaya Y, Pchelina S, Konevega A. Cryo-electron microscopy of extracellular vesicles from cerebrospinal fluid. PLoS One. 2020;15(1):e0227949. Epub 2020 Jan 30 PubMed.
Fowler SL, Behr TS, Turkes E, Cauhy PM, Foiani MS, Schaler A, Crowley G, Bez S, Ficulle E, Tsefou E, O'Brien DP, Fischer R, Geary B, Gaur P, Miller C, D'Acunzo P, Levy E, Duff KE, Ryskeldi-Falcon B. Tau filaments are tethered within brain extracellular vesicles in Alzheimer's disease. bioRxiv. 2023 Apr 30; PubMed.
Northwestern University, Feinberg School of Medicine
This interesting and rigorous study from the Frank and Ranson labs provides additional structural and molecular information on the diversity of amyloid plaques isolated from fresh App KINL-G-F brains. The author's integrated cryogenic correlated light electron microscopy and cryo-electron tomography-based analysis of hydrated plaques isolated from brain tissues nicely complements and extends previously reported amyloid structures. Maybe most interesting is their finding that the NL-G-F structures did not show an enrichment in short protofibrils as one might have expected based on results using synthetic or recombinant peptides.
Furthermore, these results provide additional evidence that Arctic Aβ fibrils differ significantly from Aβ fibrils formed in sporadic AD brain and provide some suggestions for how these structures may physically interact with additional macromolecular complexes within the extracellular space.
Florey Instotute of Neurosciece and Mental Health
University of Melbourne
Fresh eyes reveal diversity of Aβ fibrillar structures in amyloid plaques.
It’s always good to see fresh eyes looking at the major AD unsolved questions: What constitutes an Aβ plaque, its central core, peripheral halo, and surrounding tau-positive reactive neurites? When the experts claim that the Aβ-PET signals lowered by anti-amyloid therapies are the result of “plaque-lowering” effects, how do we know that all constituents of the plaque are captured by the PET-ligand in the 30 minutes after infusion? Does extra-cellular Aβ ever come into direct contact with intracellular tau? Does interaction (direct or indirect) of tau and Aβ involve liquid-liquid phase condensations, which result in various structural forms of globular/fibrillar aggregates?
These and other questions are beginning to be addressed by studies, such as this one from Leistner and colleagues in Leeds, which once again demonstrates the structural diversity of Aβ fibrillar aggregates, which are associated here with the "Arctic" Aβ /APP mutation found in familial forms of Alzheimer's disease. Notably, this mutation has led to the development of highly promising therapeutics for AD treatment, such as the FDA-approved antibody lecanemab. The cryo-EM structure of “ex vivo” purified fibrils in this study differs from previously reported structures, highlighting the notable impact of the Arctic mutation. Furthermore, these structural findings have unveiled additional fibrillar species, such as thin protofilament-like rods and branched fibrils. Nevertheless, all of these species share a common structural core element with an S-shape (Yang et al., 2022), encompassing residues 20–36 of the Aβ 42 sequence.
While cryogenic correlated light and electron microscopy (cryo-CLEM) and cryo-electron tomography (cryo-ET) have revealed that “in-tissue” Aβ fibrils are organized in a lattice or parallel bundles, and are interdigitated by subcellular compartments, extracellular vesicles, extracellular droplets, and extracellular multilamellar bodies, no information has been provided regarding the potential presence of neurofibrillary tangles (NFT) formed by tau filaments or the presence of tau filaments in sarkosyl extracts. In this study, it was assumed that AD pathologic changes include only intracellular tangles of tau. However, it has been reported that tau can be secreted into the extracellular compartment through vesicular pathways, such as exosomes, and via direct crossing of the plasma membrane. Furthermore, its interaction with lipid membranes may play a role in the formation and spreading of these pathological aggregates (El Mammeri et al., 2023). Additionally, Aβ and tau may form heterogeneous globular aggregates (Mukherjee et al., 2023), possibly through biomolecular condensation pathways and extracellular droplets, which were noted in the reported cryo-ET survey.
The identification of Aβ fibrillar deposits in the tissue was accomplished using Methoxy-X04, a Congo Red derivative known to bind to any β-pleated sheet, and which is also utilized for NFT detection (Kuchibhotla et al., 2014). Uncertainties regarding the observed fibrillar/filamentous aggregates could be addressed through immunolabeling EM, particularly by employing lecanemab. It remains unclear whether the observed “in-tissue” molecular architecture includes exclusively Aβ fibrillar forms or if other β-sheet-rich species (such as tau) or globular (co)-aggregates contribute to the lesions of AD. Finally, it might be more fruitful to shift the focus away from the use of end-stage AD brain models and place greater emphasis on investigating the earliest potential sites of Aβ/tau interaction in the precuneus in the default mode network, where the process of AD usually starts (Ruwanpathirana et al., 2022).
References:
El Mammeri N, Gampp O, Duan P, Hong M. Membrane-induced tau amyloid fibrils. Commun Biol. 2023 Apr 28;6(1):467. PubMed.
Kuchibhotla KV, Wegmann S, Kopeikina KJ, Hawkes J, Rudinskiy N, Andermann ML, Spires-Jones TL, Bacskai BJ, Hyman BT. Neurofibrillary tangle-bearing neurons are functionally integrated in cortical circuits in vivo. Proc Natl Acad Sci U S A. 2014 Jan 7;111(1):510-4. Epub 2013 Dec 24 PubMed.
Mukherjee S, Dubois C, Perez K, Varghese S, Birchall IE, Leckey M, Davydova N, McLean C, Nisbet RM, Roberts BR, Li QX, Masters CL, Streltsov VA. Quantitative proteomics of tau and Aβ in detergent fractions from Alzheimer's disease brains. J Neurochem. 2023 Feb;164(4):529-552. Epub 2022 Nov 22 PubMed.
Ruwanpathirana GP, Williams RC, Masters CL, Rowe CC, Johnston LA, Davey CE. Mapping the association between tau-PET and Aβ-amyloid-PET using deep learning. Sci Rep. 2022 Aug 30;12(1):14797. PubMed.
Yang Y, Arseni D, Zhang W, Huang M, Lövestam S, Schweighauser M, Kotecha A, Murzin AG, Peak-Chew SY, Macdonald J, Lavenir I, Garringer HJ, Gelpi E, Newell KL, Kovacs GG, Vidal R, Ghetti B, Ryskeldi-Falcon B, Scheres SH, Goedert M. Cryo-EM structures of amyloid-β 42 filaments from human brains. Science. 2022 Jan 14;375(6577):167-172. Epub 2022 Jan 13 PubMed.
Lund University
I had initially shied away from commenting mainly because the EM images with CLEM are more difficult for me to interpret. However, this is an interesting paper, and the news story and comments have added to this. I’m also prompted to write because of the suggestion in a comment to do immuno-EM with lecanemab. I agree with this suggestion and point out that we had done immuno-EM with an antibody against high-molecular-weight oligomers in AD transgenic and human biopsy brain tissue quite some time ago (Takahashi et al., 2004). Interestingly, we did see organelles that seem to be in a transition from intra- to extracellular that associated with pathological Aβ oligomer accumulations within neuritic dystrophies and with fibrillar extracellular amyloid.
Neuritic dystrophies contain the most abundant autophagic vesicles associated with plaques and could therefore be the source of the extracellular vesicles within plaques shown in this paper. However, I wonder how certain the authors are about the plasma membrane demarcations; might some amyloid fibrils intermingling with the "extracellular vesicles” possibly even be within microglia?
References:
Takahashi RH, Almeida CG, Kearney PF, Yu F, Lin MT, Milner TA, Gouras GK. Oligomerization of Alzheimer's beta-amyloid within processes and synapses of cultured neurons and brain. J Neurosci. 2004 Apr 7;24(14):3592-9. PubMed.
UCLA-DOE Molecular Biology Institute
It is amazing to be able to visualize the organization of Aβ42 fibrils in the brain of an AD model. Others have imaged amyloid fibrils in cultured cells before (Bäuerlein et al., 2017); however, this is the first time I know of that amyloid fibrils were imaged in a brain at this resolution. I am hopeful that one day it will be possible to acquire analogous images from human AD patients.
Leistner et al. showed us that the atomic-level structure of the Aβ42 fibrils purified from the mouse AD model differ from those purified from human AD patients (i.e., distinct fibril polymorphs). I am curious whether these atomic-level differences would lead to structural differences on a larger scale. That is, would the pattern of distribution of fibrils in the brains of humans with AD differ from the pattern revealed in mice by Leistner et al.? If different, it may suggest a mechanism by which fibril polymorphs define disease (viewing mouse and human AD as similar, but non-identical diseases). If the fibril distributions in human and mouse are the same, then the result would further validate the mouse system as a model for human AD.
It might be helpful, as a reference for amyloid researchers, to include cryoET density in the Amyloid Atlas [added by editors: see May 2023 news].
References:
Bäuerlein FJ, Saha I, Mishra A, Kalemanov M, Martínez-Sánchez A, Klein R, Dudanova I, Hipp MS, Hartl FU, Baumeister W, Fernández-Busnadiego R. In Situ Architecture and Cellular Interactions of PolyQ Inclusions. Cell. 2017 Sep 21;171(1):179-187.e10. Epub 2017 Sep 7 PubMed.
Make a Comment
To make a comment you must login or register.