C1q from Microglia Meddles in Neuronal Translation
Quick Links
The innate immune protein C1q seems to have a thing for neurons. Already implicated in synaptic pruning by microglia, now it is reported to also slow down protein production in neurons of the aging mouse brain. In the June 24 Cell, researchers led by Nicole Scott-Hewitt and Beth Stevens at Boston Children’s Hospital reported that microglial C1q infiltrated neuronal ribosomes in year-old mice. In vitro, C1q and RNA formed liquid droplets, hinting that C1q sequesters transcripts in the brain in a process known as liquid-liquid phase separation. In a sign of this, year-old mice that lack C1q cranked out new proteins faster than did wild-type mice.
- C1q from microglia can worm its way into the ribosomes of neurons.
- C1q binds mRNA to form liquid droplets, implying it sequesters transcripts.
- If mice lack C1q, their neurons make more protein.
“It was surprising to see C1q have such a profound effect on neuronal function,” Scott-Hewitt told Alzforum. She believes C1q may regulate translation in neurons of younger mice, but with age, these interactions turn more gel-like, clamping down on translation too much.
Other researchers praised the work. “This is an incredibly well-done and compelling study that convincingly demonstrates that a microglial-derived protein is taken up by neurons and modulates their function,” Kim Green at the University of California, Irvine, wrote to Alzforum. Borislav Dejanovic at the biotech Vigil Neuroscience in Watertown, Massachusetts, called the findings fascinating and unexpected. Because C1q is elevated in Alzheimer’s brain, its slowing of protein translation might contribute to neuronal dysfunction in that disease, Dejanovic speculated (comments below).
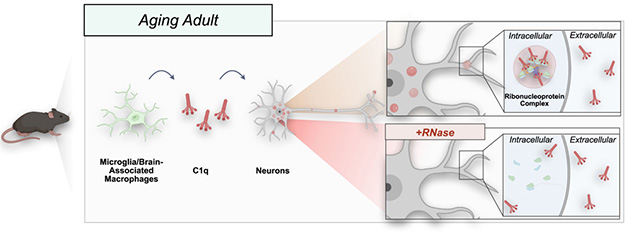
New Role for C1q. In aging mice, microglial C1q (red) gets into neurons, where it infiltrates ribosomes (top inset) and slows protein translation. Immunostaining conditions that include RNase (bottom inset) abolish this signal, showing that C1q’s association with ribosomes depends on RNA. [Courtesy of Scott-Hewitt et al., Cell.]
Stevens and others previously reported that C1q, in conjunction with other complement proteins such as C3, tags synapses for elimination during early brain development, and that this process is aberrantly reactivated in neurodegenerative disease models (Dec 2007 news; Aug 2013 news; Nov 2015 news). However, C1q also seemed to have effects that were independent of other complement proteins. Its expression climbs with age in both mouse and human brain, while those of other complement proteins do not. C1q knockout mice maintain sharper memories than wild-types as they age, and this is unrelated to C3 or synapse loss (Stephan et al., 2013). At the same time, C1q decorates synapses in the cortices of aged monkeys (Datta et al., 2020). What is it doing there?
To investigate, first author Scott-Hewitt isolated synaptosomes from wild-type mice and immunoprecipitated C1q from them, checking to see what else came along for the ride. Unexpectedly, C1q from year-old, but not 2-month-old mice, bound many ribosomal and RNA-binding proteins, suggesting the complement factor had taken up residence in neuronal ribosomes. Neurons rely on local translation at synapses to rapidly respond to stimuli.
The data puzzled Scott-Hewitt, because microglia make most of the C1q in the brain, and previous studies had detected little of it in neurons (Fonseca et al., 2017). To better visualize C1q in mouse brain sections, the authors fiddled with the immunostaining protocol, shortening the fixation time and replacing serum as a blocker of nonspecific binding with bovine serum albumin (BSA). Lo and behold, the new protocol revealed dense, punctate C1q staining throughout neuronal cell bodies and dendrites.
What explains the difference in staining? Further investigation implicated serum RNases in hiding the neuronal signal. Adding RNase to the BSA staining solution caused neuronal C1q to vanish, while adding an RNase inhibitor restored the signal even in the presence of serum. C1q’s interaction with ribosomes requires RNA, the authors concluded (image below).
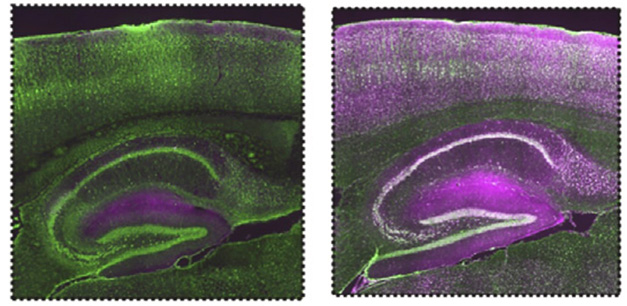
Neuronal C1q Revealed. Under typical conditions using goat serum as a blocking agent (left), immunostaining for microglial C1q (purple) shows nothing in hippocampal or cortical neurons (green). When serum is replaced with BSA (right), C1q staining pops up throughout neurons. [Courtesy of Scott-Hewitt et al., Cell.]
Commenters were impressed by this methodology. “The technical optimization … is very elegant and could be potentially of great value for the field,” Nicola Fattorelli and Renzo Mancuso at VIB-Center for Molecular Neurology, Antwerp, Belgium, wrote to Alzforum (comment below).
Delving deeper, the authors mixed C1q and total RNA in vitro. The combo phase-separated, forming droplets that grew and fused over time. This happened with human C1q and human total RNA, as well, and with RNA preparations enriched for mRNA. C1q is made up of six heterotrimers, each with a stalk and a globular head. These bind via a collagen-like domain, forming one molecule with a thicker stalk and six globular heads (image below). Because the stalk contains an intrinsically disordered region predicted to be susceptible to liquid-liquid phase separation (LLPS), the authors cleaved off this portion before combining it with RNA. The truncated C1q protein no longer formed droplets, confirming that the stalk was required for LLPS.

RNA Stalker. C1q’s stalk contains an intrinsically disordered region that can bind RNA and trigger liquid-liquid phase separation. [Courtesy of Scott-Hewitt et al., Cell.]
Given this interaction with RNA and RNA-binding proteins, might C1q affect protein translation? To test this, the authors tallied translation in wild-type and C1q knockout mice of different ages. Neurons in newborn and young adult knockouts churned out protein at the same rate as wild-types. By one year of age, however, C1q knockouts made about 30 percent more proteins than did wild types, and the mix was different.
For example, C1q knockouts upped production of mitochondrial proteins, at the expense of septins. Mitochondria are needed at synapses to provide energy for local translation. Meanwhile, septins participate in axon growth, spine formation, and synaptic vesicle release. Notably, both septins and C1q accumulate at synapses in mouse models of amyloidosis, while mitochondria are scarce (Györffy et al., 2020). Overall, the data strengthen the idea that removing C1q helps oppose Alzheimer’s-related changes at synapses.
When Scott-Hewitt and colleagues examined behavioral effects, they found that 6-week-old C1q knockouts remembered a tone associated with an electric shock for two days longer than did wild-types. It is unclear if this memory persistence in young mice is related to the enhanced memory of aging C1q knockouts.
Andrea Tenner at UC-Irvine noted that it will be important to repeat these behavioral experiments in older mice, when there is more C1q, and more droplets, in the brain. “Whether [C1q’s] RNA/RNP association has a positive or negative role in cognitive decline in disorders such as Alzheimer’s disease remains to be determined,” she wrote to Alzforum (comment below).
Serena Carra at the University of Modena and Reggio Emilia, Italy, asked whether C1q might interact with the RNA-binding protein TDP-43, which has also been implicated in suppressing protein production via LLPS (Feb 2017 news; Apr 2022 news). She also wondered which neurons are affected. “Future studies should address the question of whether C1q is preferentially taken up by specific neuronal populations, and to what extent this may contribute to the spread of pathology,” Carra wrote (comment below).—Madolyn Bowman Rogers
References
News Citations
- Paper Alert: Does the Complement Devour Synapses?
- Curbing Innate Immunity Boosts Synapses, Cognition
- Microglia Control Synapse Number in Multiple Disease States
- In New Role for TDP-43, Scientists Say it Controls Protein Synthesis
- Death by Goo: TDP-43 Gels Paralyze Proteasomes in Neurons
Paper Citations
- Stephan AH, Madison DV, Mateos JM, Fraser DA, Lovelett EA, Coutellier L, Kim L, Tsai HH, Huang EJ, Rowitch DH, Berns DS, Tenner AJ, Shamloo M, Barres BA. A Dramatic Increase of C1q Protein in the CNS during Normal Aging. J Neurosci. 2013 Aug 14;33(33):13460-74. PubMed.
- Fonseca MI, Chu SH, Hernandez MX, Fang MJ, Modarresi L, Selvan P, MacGregor GR, Tenner AJ. Cell-specific deletion of C1qa identifies microglia as the dominant source of C1q in mouse brain. J Neuroinflammation. 2017 Mar 6;14(1):48. PubMed.
- Györffy BA, Tóth V, Török G, Gulyássy P, Kovács RÁ, Vadászi H, Micsonai A, Tóth ME, Sántha M, Homolya L, Drahos L, Juhász G, Kékesi KA, Kardos J. Synaptic mitochondrial dysfunction and septin accumulation are linked to complement-mediated synapse loss in an Alzheimer's disease animal model. Cell Mol Life Sci. 2020 Feb 7; PubMed.
Other Citations
Further Reading
News
- Death by Goo: TDP-43 Gels Paralyze Proteasomes in Neurons
- Liquid Phase Transition: A Deluge of Data Points to Multiple Regulators
- How Does a Neuron Avoid Aggregation of Liquid Protein Droplets?
- Out of Chaos, Order: Reversible Amyloid Structure Seen in Phase Separation
- Tau Hooks Up with RNA to Form Droplets
- Do Membraneless Organelles Host Fibril Nucleation?
Primary Papers
- Scott-Hewitt N, Mahoney M, Huang Y, Korte N, Yvanka de Soysa T, Wilton DK, Knorr E, Mastro K, Chang A, Zhang A, Melville D, Schenone M, Hartigan C, Stevens B. Microglial-derived C1q integrates into neuronal ribonucleoprotein complexes and impacts protein homeostasis in the aging brain. Cell. 2024 Aug 8;187(16):4193-4212.e24. Epub 2024 Jun 27 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.