Microglia Make Tau, Spelling Their Demise in Tauopathies
Quick Links
In tauopathies, scientists tend to focus on neuronal tau. They track its pathology and blame it for dysfunctional microglia. Now, researchers led by Celeste Karch at Washington University, St. Louis, claim that microglia express their own tau—and that doing so does them in. In a preprint uploaded to medRxiv on May 16, they reported that iPSC-derived microglia with the IVS10+16 tau variant were sluggish at taking up myelin and tau fibrils, and at TREM2 signaling. To boot, they secreted neurotoxic molecules. Microglia isolated from human tauopathy brains also expressed tau, and their transcriptomes and proteomes suggested pathways gone awry, just as in the iMGLs.
- Microglia isolated from human brain express tau.
- Induced microglia carrying the IVS10+16 tau variant make little TREM2.
- They hardly phagocytose tau fibrils and release neurotoxic molecules.
“These data support the idea that MAPT IVS10+16 mutation may alter microglia to be less active and impaired for phagocytosis,” wrote Tsuneya Ikezu of the Mayo Clinic in Jacksonville, Florida (comment below).
Kristine Freude of University of Copenhagen, Denmark, found the evidence compelling. “This … is significant as it corroborates the limited existing studies that report tau in glial cells,” she wrote (comment below).
While highly expressed in neurons, the tau gene tends to lie fallow in other brain cells. In tauopathies, however, tau inclusions form within astrocytes and oligodendrocytes, though these likely arise from phagocytosis of tau released from neurons (Forrest et al., 2022; reviewed by Kovacs et al., 2016; Mothes et al., 2023; Ferrer et al., 2019). Microglia can adopt an inflammatory posture before tangles can even be detected in tauopathies, but again, this has been attributed to neuronal tau (Bevan-Jones et al., 2019; Bolós et al., 2015).
Karch and colleagues had another thought. What if microglia messed themselves up by expressing their own pathogenic tau? To see if this is possible, first author Abhirami Iyer analyzed previously published transcriptomic data on microglia isolated from cortical tissue of four people who had had Alzheimer’s disease and six controls from the ROSMAP cohort (Dec 2020 news). Iyer found that microglia from all donors expressed tau, though at levels much lower than what is seen in neurons.
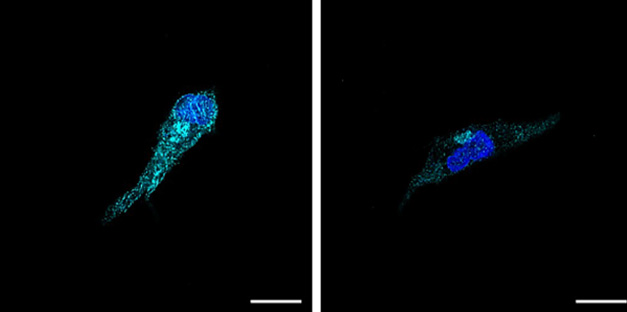
Tau Tempers TREM2. Wild type, induced microglia-like cells (left) produce much more TREM2 (cyan) than iMGLs expressing the IVS10+16 tau mutation (right). The latter have much less TREM2 on the cell surface (not shown). [Courtesy of Iyer et al., medRxiv, 2024.]
Nevertheless, the scientists studied the potential effects of this tau by creating induced microglia-like cells, i.e., iMGLs, from iPSCs of two people who each carried one copy of the MAPT IVS10+16 variant, which causes autosomal-dominant frontotemporal dementia. Like microglia in vivo, iMGLs expressed tau mRNA and protein. The IVS10+16 iMGLs made as much tau as isogenic control cells, though they favored four-repeat over three-repeat tau, in keeping with the variant’s ability to alter tau splicing in neurons (Capano et al., 2022).
RNA-Seq revealed that the transcriptional states had shifted in IVS10+16 iMGLs. They mobilized chemokine genes at the expense of genes needed for phagocytosis, or of genes from disease-associated and lipid-associated microglia of AD and other neurodegenerative diseases express. As a result, the tau variant iMGLs engulfed about one-third less fibrillar tau and 23 percent less myelin than did control cells, highlighting their listless phagocytosis.
The downregulation of DAM and LAM genes hinted that the IVS10+16 iMGLs might be less reactive than control cells. They had little TREM2 mRNA and almost none of the receptor protein. A DAM gene, TREM2 spurs microglia to engulf myelin debris as well as surround and compact amyloid plaques (see image above).

Totally Tubulin. Rather than branching out from the nucleus, as in wild-type iMGLs (left), α-tubulin (black) gathers around the cell periphery in IVS10+16 iMGLs (right). [Courtesy of Iyer et al., medRxiv, 2024.]
Given that tau binds microtubules, the scientists next looked at the iMGL cytoskeleton. The mutant tau cells had quieted genes regulating actin, a microfilament building block, and had only half as much actin as isogenic controls. The microtubule protein α-tubulin was distributed abnormally within the induced microglia, hanging around the furthest reaches of the cytosol rather than near the nucleus (image at right). The authors believe wonky cytoskeletons explain the phagocytosis defect, since that process requires the cytoskeleton to wrap around fragments of myelin, tau, and other debris.
To learn if IVS10+16 tau affected the microglial secretome, Iyer measured levels of 1,360 proteins in the iMGLs culture media. Eighty-nine were differently expressed in IVS10+16 versus control iMGLs, with most being overproduced (image below). The scientists think these 89 make for a toxic brew, since medium from tau-variant iMGLs stunted synapses when added to iPSC-derived neurons in culture.

Noxious Secretome? IVS10+16 iMGLs overproduced most secretory proteins (red), but held others back (blue). [Courtesy of Iyer et al., medRxiv, 2024.]
All told, the findings indicate that mutant tau has profound cell-autonomous effects on induced microglia. Would that play out in vivo? Many scientists do not believe induced microglia fully recapitulate the properties of microglia in their normal environment. In fact, Kiran Bhaskar, Jonathan Hulse, and Karthikeyan Tangavelou, all at the University of New Mexico in Albuquerque, wondered if the iMGLs turned on neuronal genes, including tau, because they were in culture (comment below).
To look for in vivo evidence of cell-autonomous effects, Iyer and colleagues combed through published bulk RNA-Seq data on middle temporal gyrus tissue from three controls and two people carrying MAPT IVS10+16. They found 100 genes whose up- or downregulation in carriers mimicked that seen in iMGLs (Minaya et al., 2023). These included immune activation and cytoskeletal organization genes. Likewise, CSF proteomics data from 37 people with various tau mutations were compatible with an uptick in microglial adaptive immunity pathways, and a failing of extracellular matrix pathways, just as in the iMGLs.
To Karch, this was profound. “I was blown away by the degree to which the proteins that changed in people with tauopathies matched those that changed in the media of the iMGLs,” she told Alzforum. To her mind, these findings highlight how accurately iMGLs reflect what is going on in people.
Karch wants to understand how these findings pertain to other tau mutations, so she is repeating the experiments using the P301L, R406W, and V337M tau mutations.—Chelsea Weidman Burke
References
News Citations
Mutations Citations
Paper Citations
- Forrest SL, Wagner S, Kim A, Kovacs GG. Association of glial tau pathology and LATE-NC in the ageing brain. Neurobiol Aging. 2022 Nov;119:77-88. Epub 2022 Jul 31 PubMed.
- Kovacs GG, Ferrer I, Grinberg LT, Alafuzoff I, Attems J, Budka H, Cairns NJ, Crary JF, Duyckaerts C, Ghetti B, Halliday GM, Ironside JW, Love S, Mackenzie IR, Munoz DG, Murray ME, Nelson PT, Takahashi H, Trojanowski JQ, Ansorge O, Arzberger T, Baborie A, Beach TG, Bieniek KF, Bigio EH, Bodi I, Dugger BN, Feany M, Gelpi E, Gentleman SM, Giaccone G, Hatanpaa KJ, Heale R, Hof PR, Hofer M, Hortobágyi T, Jellinger K, Jicha GA, Ince P, Kofler J, Kövari E, Kril JJ, Mann DM, Matej R, McKee AC, McLean C, Milenkovic I, Montine TJ, Murayama S, Lee EB, Rahimi J, Rodriguez RD, Rozemüller A, Schneider JA, Schultz C, Seeley W, Seilhean D, Smith C, Tagliavini F, Takao M, Thal DR, Toledo JB, Tolnay M, Troncoso JC, Vinters HV, Weis S, Wharton SB, White CL 3rd, Wisniewski T, Woulfe JM, Yamada M, Dickson DW. Aging-related tau astrogliopathy (ARTAG): harmonized evaluation strategy. Acta Neuropathol. 2016 Jan;131(1):87-102. Epub 2015 Dec 10 PubMed.
- Mothes T, Portal B, Konstantinidis E, Eltom K, Libard S, Streubel-Gallasch L, Ingelsson M, Rostami J, Lindskog M, Erlandsson A. Astrocytic uptake of neuronal corpses promotes cell-to-cell spreading of tau pathology. Acta Neuropathol Commun. 2023 Jun 17;11(1):97. PubMed.
- Ferrer I, Aguiló García M, Carmona M, Andrés-Benito P, Torrejón-Escribano B, Garcia-Esparcia P, Del Rio JA. Involvement of Oligodendrocytes in Tau Seeding and Spreading in Tauopathies. Front Aging Neurosci. 2019;11:112. Epub 2019 May 28 PubMed.
- Bevan-Jones WR, Cope TE, Jones PS, Passamonti L, Hong YT, Fryer T, Arnold R, Coles JP, Aigbirhio FI, O'Brien JT, Rowe JB. In vivo evidence for pre-symptomatic neuroinflammation in a MAPT mutation carrier. Ann Clin Transl Neurol. 2019 Feb;6(2):373-378. Epub 2019 Jan 2 PubMed.
- Bolós M, Llorens-Martín M, Jurado-Arjona J, Hernández F, Rábano A, Avila J. Direct Evidence of Internalization of Tau by Microglia In Vitro and In Vivo. J Alzheimers Dis. 2015;50(1):77-87. PubMed.
- Capano LS, Sato C, Ficulle E, Yu A, Horie K, Kwon JS, Burbach KF, Barthélemy NR, Fox SG, Karch CM, Bateman RJ, Houlden H, Morimoto RI, Holtzman DM, Duff KE, Yoo AS. Recapitulation of endogenous 4R tau expression and formation of insoluble tau in directly reprogrammed human neurons. Cell Stem Cell. 2022 Jun 2;29(6):918-932.e8. PubMed.
- Minaya MA, Mahali S, Iyer AK, Eteleeb AM, Martinez R, Huang G, Budde J, Temple S, Nana AL, Seeley WW, Spina S, Grinberg LT, Harari O, Karch CM. Conserved gene signatures shared among MAPT mutations reveal defects in calcium signaling. Front Mol Biosci. 2023;10:1051494. Epub 2023 Feb 9 PubMed.
Further Reading
No Available Further Reading
Primary Papers
- Iyer AK, Vermunt L, Mirfakhar FS, Minaya M, Acquarone M, Koppisetti RK, Renganathan A, You S-F, Danhash E, Verbeck A, Galasso G, Lee SM, Marsh J, Nana AL, Spina S, Seeley WW, Grinberg LT, Temple S, Teunissen CE, Sato C, Karch C. Cell autonomous microglia defects in a stem cell model of frontotemporal dementia. 2024 May 16 10.1101/2024.05.15.24307444 (version 1) medRxiv.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.