Live-Cell Studies Blame Arginine Peptides for C9ORF72’s Crimes
Quick Links
A variety of abnormal RNAs and peptides result from repeats in the C9ORF72 gene, the most common genetic cause of frontotemporal dementia and amyotrophic lateral sclerosis. Which of these funky molecules instigates the death of cortical and motor neurons? Researchers from Thomas Jefferson University in Philadelphia have used live-cell imaging to catch the true killers red-handed. Their work points the finger at both the guanine- and cytosine-rich repeat RNA as well as arginine-rich peptides, particularly the proline-arginine version. The RNAs and peptides conspire to infiltrate the nucleolus, causing rapid neuron death, report senior author Davide Trotti and colleagues in the December 17 Neuron. They also demonstrate, for the first time, that the arginine-containing peptides appear in cell lines derived from the tissues of people with C9ORF72 repeats.
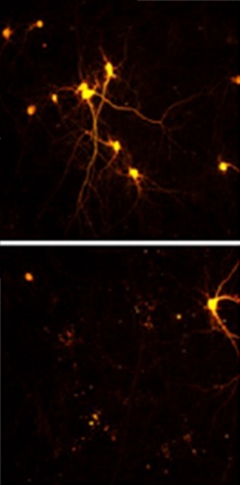
Deadly Dipeptides:
Cortical neurons normally survive in culture (top), but those expressing the PR dipeptide disappear within eight days (bottom). [Image courtesy of Neuron, Wen et al.]
Most people have a handful of GGGGCC repeats in the C9ORF72 gene, but the hexanucleotide can number hundreds or thousands in people with ALS or FTD (see Sep 2011 news story). Researchers were surprised to find that despite being in an intron, the repeats are transcribed into RNAs that go in both the sense and antisense directions. The six reading frames translate into five possible repeat dipeptides: proline-arginine, proline-alanine, glycine-alanine, glycine-arginine, and glycine-proline (see Nov 2013 news story). Some or all of these molecules might instigate neurodegeneration. Alternatively, researchers have hypothesized that the disease results from what is missing: Repeats interfere with production of normal C9ORF72, and the cells could die from dearth of the protein (Ciura et al., 2013).
To sift through these hypotheses, first author Xinmei Wen opted to study cell types as similar as possible to the motor neurons and cortical neurons that degenerate in ALS and FTD, respectively. Previous studies had focused on neuroblastoma cell lines or non-neuronal cells, such as immortalized bone and kidney cells. Instead, Wen and colleagues used primary cortical and motor neurons from embryonic rats. They also derived motor neurons from induced pluripotent stem cells (iPSCs) generated from people with C9ORF72 expansions. Wen engineered cells to produce the dipeptides only (i.e., no RNA), express the repeat RNA only (no peptide), or make neither (C9ORF72 knockdown). The researchers used an automated microscope to check on individual cell survival every 24 hours.
First up, repeat dipeptides. Wen transfected the primary cells with DNA that would make each peptide with 50 repeats. She swapped out the GC-rich codons for others that corresponded to the same amino acids, so the cells would churn out peptides without making RNA containing GGGGCC repeats. She tagged the peptides with green fluorescent protein. Among cells producing proline-alanine, glycine-alanine, or glycine-proline, at least half survived for five days or more in culture. Proline-arginine, in contrast, killed half of the cells within three days. Glycine-arginine exhibited intermediate toxicity, killing half the motor neurons by four days but less than a third of the cortical neurons. The arginine-containing dipeptides are the “lethal” agent, Trotti concluded.
What makes proline-arginine and glycine-arginine so nasty? Using computer algorithms to analyze the sequence, Wen noticed that the heavily basic arginine-containing sequences mimic nucleolar localization signals. Indeed, the green fluorescent protein signal showed that proline-arginine and glycine-arginine aggregated in this organelle, while the others remained cytoplasmic. While glycine-arginine sometimes dawdled in the cytoplasm, proline-arginine always went straight for the nucleolus, perhaps explaining why it was most toxic, Trotti said. In separate experiments, aggregation of proline-arginine led to the formation of stress granules in the cytoplasm. These RNA-protein globs, which preserve transcripts during times of cellular stress, have been linked to neurodegeneration (reviewed in Wolozin, 2014).
Next, Wen turned her attention to the GGGGCC RNA. Previous work indicated that these form disruptive aggregates (see Oct 2013 news story). To separate the effects of the RNA from the peptides it encodes, Wen engineered DNA encoding 42 hexanucleotide repeats, but in an intron she added to the green fluorescent protein gene. She confirmed by western blotting that these repeats were not translated into peptides. When Wen transfected this construct into cortical neurons it was toxic, but did not make as big a difference to cell survival as the arginine-rich peptides. Therefore, she used the live-cell imaging over time to calculate the risk of death for cells in the culture. Cells with the repeat RNA were about twice as likely to die as control neurons.
If both RNA and dipeptides are toxic, might they work in concert? Wen claimed they have a synergistic effect because the risk of cell death was greater for cells transfected with both proline and hexanucleotide repeat constructs than when transfected with either alone. The authors concluded that the GGGGCC RNA and proline-arginine dipeptide work together in the same toxic pathway.
Finally, Wen addressed the hypothesis that neurons need C9ORF72 to survive. She used interfering RNA to knock down the gene’s expression by about 70 percent. This did not influence motor neuron survival. However, Wen pointed out that this does not completely refute the haploinsufficiency hypothesis; lowered production of C9ORF72 over a lifetime might cause disease, even though isolated neurons can live for days with subnormal expression.
Since the rat neuron experiments pointed to dipeptides as the most toxic entity, Wen wanted to find evidence of toxic proline-arginine in human cells. She induced motor neurons from IPSCs derived from people with C9ORF72 expansions. Cultures of these neurons accumulated proline-arginine aggregates, both in the nucleus and outside of the cells, stuck to the bottom of the dish. The extracellular aggregates may have been left over from neurons that died, Trotti suggested. Wen also found proline-arginine aggregates in spinal cord tissue from people who died of C9ORF72-related ALS.
Wen’s work falls in line with studies that suggest that arginine-rich peptides are the worst offenders in C9ORF72 disease (see Aug 2014 news story; news story), but conflicts with others that finger the glycine-alanine dipeptide (May et al., 2014; Zhang et al., 2014). Though glycine-arginine and proline-arginine are clearly toxic in Wen’s and others’ experimental models, Dieter Edbauer of Ludwig-Maximilians University in Munich wrote that he still leans toward glycine-alanine as the primary troublemaker in people (see full comment below). In patient brains, the arginine-rich peptides tend to accumulate in the cytoplasm rather than nucleus, he explained, which means cells may not be good models of the human condition. Plus, glycine-alanine co-aggregates with p62 and ubiquitin, typical markers of ALS pathology, while glycine-arginine and proline-arginine do not (Yamakawa et al., 2014).
Wen’s work adds to several studies examining C9ORF72 mechanisms in vitro. In the future, it will be exciting for researchers to also determine how much the arginine-rich dipeptides contribute to disease in people, pointed out Jiou Wang of Johns Hopkins University in Baltimore, who was not involved in this publication. That will help in developing therapies to block the dipeptide’s dirty work, he said.—Amber Dance
References
News Citations
- Corrupt Code: DNA Repeats Are Common Cause for ALS and FTD
- Sense, Antisense: C9ORF72 Makes Both Forms of RNA, Peptides
- RNA Deposits Confer Toxicity in C9ORF72 ALS
- C9ORF72 Killer Dipeptides Clog the Nucleolus
- C9ORF72’s Dirty Work Done by Problem Proteins
Paper Citations
- Ciura S, Lattante S, Le Ber I, Latouche M, Tostivint H, Brice A, Kabashi E. Loss of function of C9orf72 causes motor deficits in a zebrafish model of Amyotrophic Lateral Sclerosis. Ann Neurol. 2013 May 30; PubMed.
- Wolozin B. Physiological protein aggregation run amuck: stress granules and the genesis of neurodegenerative disease. Discov Med. 2014 Jan;17(91):47-52. PubMed.
- May S, Hornburg D, Schludi MH, Arzberger T, Rentzsch K, Schwenk BM, Grässer FA, Mori K, Kremmer E, Banzhaf-Strathmann J, Mann M, Meissner F, Edbauer D. C9orf72 FTLD/ALS-associated Gly-Ala dipeptide repeat proteins cause neuronal toxicity and Unc119 sequestration. Acta Neuropathol. 2014 Oct;128(4):485-503. Epub 2014 Aug 14 PubMed.
- Zhang YJ, Jansen-West K, Xu YF, Gendron TF, Bieniek KF, Lin WL, Sasaguri H, Caulfield T, Hubbard J, Daughrity L, Chew J, Belzil VV, Prudencio M, Stankowski JN, Castanedes-Casey M, Whitelaw E, Ash PE, DeTure M, Rademakers R, Boylan KB, Dickson DW, Petrucelli L. Aggregation-prone c9FTD/ALS poly(GA) RAN-translated proteins cause neurotoxicity by inducing ER stress. Acta Neuropathol. 2014 Oct;128(4):505-24. Epub 2014 Aug 31 PubMed.
- Yamakawa M, Ito D, Honda T, Kubo K, Noda M, Nakajima K, Suzuki N. Characterization of the dipeptide repeat protein in the molecular pathogenesis of c9FTD/ALS. Hum Mol Genet. 2015 Mar 15;24(6):1630-45. Epub 2014 Nov 14 PubMed.
Further Reading
Papers
- Liu EY, Russ J, Wu K, Neal D, Suh E, McNally AG, Irwin DJ, Van Deerlin VM, Lee EB. C9orf72 hypermethylation protects against repeat expansion-associated pathology in ALS/FTD. Acta Neuropathol. 2014 Oct;128(4):525-41. Epub 2014 May 8 PubMed.
- Waite AJ, Bäumer D, East S, Neal J, Morris HR, Ansorge O, Blake DJ. Reduced C9orf72 protein levels in frontal cortex of amyotrophic lateral sclerosis and frontotemporal degeneration brain with the C9ORF72 hexanucleotide repeat expansion. Neurobiol Aging. 2014 Jul;35(7):1779.e5-1779.e13. Epub 2014 Jan 17 PubMed.
- Proudfoot M, Gutowski NJ, Edbauer D, Hilton DA, Stephens M, Rankin J, Mackenzie IR. Early dipeptide repeat pathology in a frontotemporal dementia kindred with C9ORF72 mutation and intellectual disability. Acta Neuropathol. 2014 Mar;127(3):451-8. Epub 2014 Jan 21 PubMed.
- Mackenzie IR, Frick P, Neumann M. The neuropathology associated with repeat expansions in the C9ORF72 gene. Acta Neuropathol. 2014 Mar;127(3):347-57. Epub 2013 Dec 20 PubMed.
- Mackenzie IR, Arzberger T, Kremmer E, Troost D, Lorenzl S, Mori K, Weng SM, Haass C, Kretzschmar HA, Edbauer D, Neumann M. Dipeptide repeat protein pathology in C9ORF72 mutation cases: clinico-pathological correlations. Acta Neuropathol. 2013 Dec;126(6):859-79. Epub 2013 Oct 6 PubMed.
Primary Papers
- Wen X, Tan W, Westergard T, Krishnamurthy K, Markandaiah SS, Shi Y, Lin S, Shneider NA, Monaghan J, Pandey UB, Pasinelli P, Ichida JK, Trotti D. Antisense proline-arginine RAN dipeptides linked to C9ORF72-ALS/FTD form toxic nuclear aggregates that initiate in vitro and in vivo neuronal death. Neuron. 2014 Dec 17;84(6):1213-25. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
DZNE
To use a sports analogy it's now 3:3—the glycine-alanine camp (May et al., 2014; Zhang YJ et al., 2014; Yamakawa M et al., 2014) and the arginine-rich camp (Kwon I et al., 2014; Mizielinska S et al., 2014; Wen X et al., 2014) are tied. I find the cell type specific toxicity and the interaction of dipeptide repeat (DPR) and RNA toxicity the most interesting results in this, the latest DPR toxicity paper from the Trotti lab. Moreover, it's nice that somebody finally published that C9ORF72 knockdown is not overtly neurotoxic, which argues against a major role for C9ORF72 loss of function in ALS/FTD.
I still favor the role of glycine-alanine toxicity for the pathogenesis, although recombinant proline-arginine and glycine-arginine clearly can induce toxicity in various systems (including some of our experiments). My main concern with PR/GR toxicity is the nucleolar aggregation pattern of recombinant PR and GR proteins in vitro, which is in contrast to the predominant cytoplasmic aggregation of PR and GR seen in patient brains. At least with our antibodies, the less common nuclear aggregates don't seem to co-localize with nucleoli. Also, unlike the DPR inclusions in patients, the aggregates of recombinant PR/GR are p62 negative. Since recombinant GA expressed in cell culture looks very similar to GA found in patient tissue, we have been focusing our analysis on GA toxicity.
I believe that when two camps are arguing in science, the truth usually lies in between. Mouse models and more detailed neuropathological analysis will be important for settling this issue. If nucleolar PR/GR is indeed the main culprit, I expect that future studies will find a better correlation between this regional neurodegeneration and this specific pathology rather than for GA aggregates. On the other hand, if cells with nucleolar PR/GR aggregates die too quickly to be found in postmortem tissue, it will be hard to show their clinical relevance other than by removing them therapeutically.
View all comments by Dieter EdbauerMake a Comment
To make a comment you must login or register.