In Frontotemporal Lobar Dementia, TDP-43 Snaps into a Chevron Shape
Quick Links
Following in the footsteps of tau and α-synuclein, TDP-43, too, appears to twist into distinct amyloid arrangements in different neurodegenerative conditions. According to a paper published in Nature on August 2, filaments of TDP-43 in brain samples from people with frontotemporal lobar degeneration (FTLD) type A share a protofilament core that folds into a chevron-like shape. This new amyloid fold markedly differs from the double spiral contortion previously found in people with a form of FTLD accompanied by amyotrophic lateral sclerosis. The authors, led by Benjamin Ryskeldi-Falcon of the MRC Laboratory of Molecular Biology in Cambridge, U.K., also zeroed in on a post-translational tweak—citrullination of an arginine residue—that may trigger its formation.
- In three people with FTLD-A, the core of TDP-43 filaments folds into a chevron pattern.
- This fold is distinct from the one TDP-43 takes in people with FTD/ALS.
- Modification of a core arginine residue may facilitate the chevron shape.
To Ryskeldi-Falcon’s mind, the protofilament folds taken by TDP-43 in different forms of FTLD support an emerging theme in neurodegenerative proteinopathies, i.e., that distinct filament structures define different diseases.
Along with filaments of TDP-43, the researchers also detected TMEM106b fibrils in the same brain samples. This corroborates three previous studies that identified TMEM106b fibrils in brain samples from people with different forms of FTLD and other neurodegenerative diseases. One group had claimed that the abundant TDP-43 aggregates in these samples were not fibrillar (Apr 2022 news on Jiang et al., 2022). Now that filaments of both proteins have been clearly identified in the same brain samples, it seems that the discrepancy likely comes down to different biochemical extraction procedures, Ryskeldi-Falcon told Alzforum.
To Christian Haass of the German Center for Neurodegenerative Diseases in Munich, the unambiguous presence of TDP-43 filaments in different pathological forms of FTLD further supports a pathological nature of these aggregates, and unveils new opportunities for therapeutic targeting (comment below).
FTLD comprises a spectrum of disorders marked by extreme heterogeneity at both the neuropathological and clinical level. About half of cases carry TDP-43 inclusions, which, depending on their morphology and distribution within the brain, are classified into four types, dubbed A-D (Neumann et al., 2021). Previously, Ryskeldi-Falcon and colleagues had resolved the structure of TDP-43 filaments in people with both FTLD/ALS who had TDP-43 type B pathology, in which moderate numbers of neuronal cytoplasmic inclusions of TDP-43 gather in both the superficial and deep layers of the cortex, in the absence of dystrophic neurites. Whether plucked from the frontal or motor cortices, this brand of TDP-43 filament contained a core that twisted into a double-spiral fold (Dec 2021 news). Would the RNA-binding protein twist into the same configuration in people with different neuropathological and clinical manifestations of this proteinopathy?
To find out, first author Diana Arseni and colleagues turned the cryo-electron microscope on filaments extracted from brain samples rife with the most common form of TDP-43 pathology, i.e., type A. Abundant neuronal cytoplasmic inclusions of the protein, which are concentrated in the superficial layers of the cortex and accompanied by short, thick dystrophic neurites, characterize this disease. Type A TDP-43 pathology mostly strikes people with FTD who do not have ALS. Brain samples were provided by Masato Hasegawa of Tokyo Metropolitan University and Bernardino Ghetti of Indiana University in Indianapolis. Three samples were used for cryo-EM, including one whose clinical presentation was behavioral variant of FTD, and two others who suffered from non-fluent variant of primary progressive aphasia. The latter two carried a progranulin mutation.
Arseni spotted TDP-43 filaments lurking in all three samples. The filaments twisted to the right along their axes. Their cores consisted of single molecules of TDP-43 stacked on top of each other. Other fibril protofilaments often consist of dimers, as in the case of tau and Aβ filaments (Jul 2017 news; Jan 2022 news).
With cryo-EM, the scientists resolved the shape of each TDP-43 molecule in this ordered core, revealing that a stretch of residues from R272 to Q360 of the low-complexity domain of TDP-43 bends and folds over itself four times to form five “layers” (image below).

Behold the Chevron Fold. In filaments from three people with type A FTLD (top), a portion of TDP-43’s low-complexity domain folds over itself to create five layers (top). A model of the protein (bottom) shows that layers 3, 4, and 5 each form a V. Together, all three form a chevron, the authors suggest. [Courtesy of Arnesi et al., Nature, 2023.]
This five-layer structure centers around a hydrophobic stretch of residues that form a kinked β-strand within the fourth layer. This central β-strand forms steric zippers with amino acids in the neighboring third and fifth layers of the fold. Each of these three zipped-up sections of the protein form a V. Together they create a shape akin to a chevron, i.e., three Vs in a row. Layers 1 and 2 at the core's N-terminus comprise shorter β-strands that do not partake in the chevron.
Residues that lie N- and C-terminal to the ordered core form an unstructured fuzzy coat around the filaments, which was not resolved by cryo-EM. In addition to unresolved segments of this coat, non-proteinaceous densities mingled between the first, second, and third layers of the protofilament core, suggesting the presence of possible co-factors.
All the TDP-43 filaments observed from the three samples contained a chevron fold at their core. However, in about 5 percent of fibrils, the chevron fold took a slightly different configuration at its N-terminus and at the turn between the third and fourth layers. Interestingly, the scientists spotted this “minority chevron” within fibrils containing the predominant fold, suggesting the two were compatible variations, rather than entirely distinct folds. The chevron fold was markedly distinct from the fold Ryskeldi-Falcon’s group had previously described in people with FTLD type B pathology, although it did involve a similar segment of the protein (image below).
“This important study now provides the direct proof for different TDP-43 conformations as the probable molecular basis for the phenotypic variability among TDP-43 proteinopathies,” wrote Manuela Neumann of the German Center for Neurodegenerative Diseases in Tübingen (comment below). Neumann discovered the TDP-43 aggregates that led to the identification of the different neuropathological subtypes of FTLD (Neumann et al., 2006).
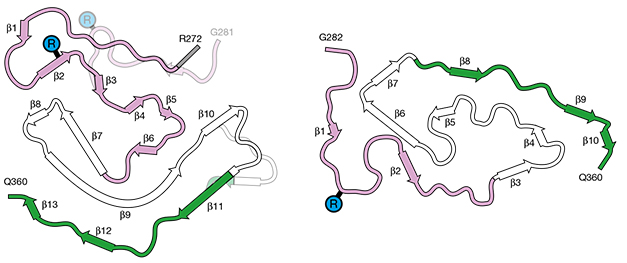
Variations on a Fold. The chevron structure (left) assumed by TDP-43 in filaments in FTLD type A differs from the spiral (right) found in FTLD type B filaments. [Courtesy of Arseni et al., Nature, 2023.]
What makes TDP-43 fold into this distinct structure? This question remains unanswered. A clue might come from R293, an arginine residue sandwiched snugly between layers 1 and 2 of the protofilament fold. With its positive charge only partially compensated for by surrounding peptide groups, this arginine should be “very unhappy” in this position, Ryskeldi-Falcon said. The only explanation for its placement? A post-translational modification that neutralizes its charge, he said. In the case of arginine, citrullination, aka deamination, would fit the bill. Indeed, mass spectrometry confirmed that R293 was citrullinated in TDP-43 from brain samples of people with FTLD type A pathology. Ryskeldi-Falcon proposed that without this modification, the chevron fold could not have formed.
Mass spec also detected some R293 residues donning a methyl group, a modification that would be expected to fit within the slightly altered chevron fold taken by a minority of protofilaments. Ryskeldi-Falcon proposed that these post-translational modifications likely govern how the fold forms, as well as its variations.
Colin Masters and Victor Streltsov of the University of Melbourne agreed. They went a step further. “This suggests a broader concept where specific disease-related, post-translational modification patterns determine the amyloid filament conformation of distinct proteins in neurodegenerative diseases,” they wrote (comment below).
Beyond this single arginine residue in TDP-43, emerging evidence suggests that citrullination could play a broader role in the function and aggregation of proteins involved in neurodegenerative disease. For example, recent studies have reported an uptick in the activity of protein arginine deaminases—the enzymes that perform citrullination—in the brains of people with different neurodegenerative conditions (Feb 2018 news; Ishigami et al., 2005; Mondal et al., 2021). Other emerging studies cast citrullination as a potential cause of harmful autoimmune reactions that underlie diseases such as multiple sclerosis (Martín Monreal et al., 2023).
Maj-Linda Selenica of the University of Kentucky in Lexington studies the role of citrullination on TDP-43 function and aggregation. She told Alzforum that her group has found 11 of the 20 TDP-43 arginine residues can become citrullinated, and the modification appears to strongly influence the protein's liquid-liquid phase separation, pushing it to condense into small droplets. Selenica’s group has generated antibodies specific for these different modified forms. Preliminary findings suggest that TDP-43 citrullination rises in people with limbic predominant TDP-43 encephalopathy (LATE), a proteinopathy that causes cognitive decline and often co-occurs with AD. Selenica was excited to see that citrullination appears to be involved in the aggregation of TDP-43 in FTD, and believes targeting the pathway could hold therapeutic promise.
In addition to TDP-43 fibrils, Arseni also detected TMEM106b fibrils lurking in all three of the brain samples. These did not co-localize with the TDP-43 filaments. Their structure was consistent with the shape previously spotted among people with FTD/ALS type B pathology, as well as among people with other neurodegenerative diseases and even those without a proteinopathy (Apr 2022 news).
Thus far, TMEM106b filament structures do not appear specific to disease type, in contrast to fibrils of TDP-43 and other proteins, including tau, and α-synuclein. “The available evidence is consistent with the age-dependent accumulation of TMEM106B filaments in the human brain,” the authors concluded. —Jessica Shugart
References
News Citations
- Surprise! TMEM106b Fibrils Found in Neurodegenerative Diseases
- Double Spiral Sets TDP-43 Apart from Other Amyloids
- Tau Filaments from the Alzheimer’s Brain Revealed at Atomic Resolution
- Cryo-EM Unveils Distinct Aβ42 Fibril Structures for Sporadic, Familial AD
- Citrullination, Anyone? New Gene Implicated in ALS
Paper Citations
- Jiang YX, Cao Q, Sawaya MR, Abskharon R, Ge P, DeTure M, Dickson DW, Fu JY, Ogorzalek Loo RR, Loo JA, Eisenberg DS. Amyloid fibrils in FTLD-TDP are composed of TMEM106B and not TDP-43. Nature. 2022 May;605(7909):304-309. Epub 2022 Mar 28 PubMed.
- Neumann M, Lee EB, Mackenzie IR. Frontotemporal Lobar Degeneration TDP-43-Immunoreactive Pathological Subtypes: Clinical and Mechanistic Significance. Adv Exp Med Biol. 2021;1281:201-217. PubMed.
- Neumann M, Sampathu DM, Kwong LK, Truax AC, Micsenyi MC, Chou TT, Bruce J, Schuck T, Grossman M, Clark CM, McCluskey LF, Miller BL, Masliah E, Mackenzie IR, Feldman H, Feiden W, Kretzschmar HA, Trojanowski JQ, Lee VM. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006 Oct 6;314(5796):130-3. PubMed.
- Ishigami A, Ohsawa T, Hiratsuka M, Taguchi H, Kobayashi S, Saito Y, Murayama S, Asaga H, Toda T, Kimura N, Maruyama N. Abnormal accumulation of citrullinated proteins catalyzed by peptidylarginine deiminase in hippocampal extracts from patients with Alzheimer's disease. J Neurosci Res. 2005 Apr 1;80(1):120-8. PubMed.
- Mondal S, Thompson PR. Chemical biology of protein citrullination by the protein A arginine deiminases. Curr Opin Chem Biol. 2021 Aug;63:19-27. Epub 2021 Mar 4 PubMed.
- Martín Monreal MT, Hansen BE, Iversen PF, Enevold C, Ødum N, Sellebjerg F, Højrup P, Rode von Essen M, Nielsen CH. Citrullination of myelin basic protein induces a Th17-cell response in healthy individuals and enhances the presentation of MBP85-99 in patients with multiple sclerosis. J Autoimmun. 2023 Jul 26;139:103092. PubMed.
Further Reading
No Available Further Reading
Primary Papers
- Arseni D, Chen R, Murzin AG, Peak-Chew SY, Garringer HJ, Newell KL, Kametani F, Robinson AC, Vidal R, Ghetti B, Hasegawa M, Ryskeldi-Falcon B. TDP-43 forms amyloid filaments with a distinct fold in type A FTLD-TDP. Nature. 2023 Aug;620(7975):898-903. Epub 2023 Aug 2 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Tübingen and DZNE AG Neumann
Benjamin Ryskeldi-Falcon, Diane Arseni, and colleagues have extended their fantastic cryo-EM work in TDP-43 proteinopathies. After reporting the first structure of TDP-43 filaments in ALS/FTLD-TDP type B cases last year (Arseni et al., 2022), in this study they now resolved the structure(s) of TDP-43 filaments in FTLD-TDP type A cases. Importantly, the filaments in FTLD-TDP type A and B are structurally different, thereby providing the first direct evidence for the presence of distinct TDP-43 conformations.
Since initial reports in 2006, subclassification of FTLD-TDP based on the pattern of neocortical TDP-43 pathology, with each subtype showing specific clinical and genetic correlations, became widely accepted in the field (Mackenzie et al., 2009; Mackenzie and Neumann, 2017).
By analogy to other neurodegenerative diseases, a popular hypothesis to explain the heterogeneity in FTLD-TDP is to propose the presence of different conformational types of misfolded TDP-43, or “TDP-43 strains”, with each strain associated with specific cell tropism, seeding activity, and toxicity. While there was already growing, indirect evidence supporting the idea of distinct conformational TDP-43 species, with demonstration of biochemical differences of insoluble TDP-43 as well as identification of type-specific TDP-43 antibodies (reviewed in Neumann et al., 2021), this important study now provides the direct proof for different TDP-43 conformations as the likely molecular basis for the phenotypic variability among TDP-43 proteinopathies. I am very much looking forward to seeing the structure of FTLD-TDP type C filaments in the near future.
References:
Arseni D, Hasegawa M, Murzin AG, Kametani F, Arai M, Yoshida M, Ryskeldi-Falcon B. Structure of pathological TDP-43 filaments from ALS with FTLD. Nature. 2022 Jan;601(7891):139-143. Epub 2021 Dec 8 PubMed.
Mackenzie IR, Neumann M, Bigio EH, Cairns NJ, Alafuzoff I, Kril J, Kovacs GG, Ghetti B, Halliday G, Holm IE, Ince PG, Kamphorst W, Revesz T, Rozemuller AJ, Kumar-Singh S, Akiyama H, Baborie A, Spina S, Dickson DW, Trojanowski JQ, Mann DM. Nomenclature for neuropathologic subtypes of frontotemporal lobar degeneration: consensus recommendations. Acta Neuropathol. 2009 Jan;117(1):15-8. PubMed.
Mackenzie IR, Neumann M. Reappraisal of TDP-43 pathology in FTLD-U subtypes. Acta Neuropathol. 2017 Jul;134(1):79-96. Epub 2017 May 2 PubMed.
Neumann M, Lee EB, Mackenzie IR. Frontotemporal Lobar Degeneration TDP-43-Immunoreactive Pathological Subtypes: Clinical and Mechanistic Significance. Adv Exp Med Biol. 2021;1281:201-217. PubMed.
University of Melbourne
Florey Instotute of Neurosciece and Mental Health
The twists and turns of TDP
The argument regarding whether the presence of specific fibrils in particular cases stems from different extraction procedures or case-specific neuropathological conditions seems to find support from both sides. This study led by Ryskeldi-Falcon describes the cryo-EM structure of TDP-43 filaments found in three patients with FTLD type A. These filaments exhibit a distinct five-layered core protofilament folded into a chevron-like shape, unlike the previously observed double spiral fold among individuals with FTLD/ALS (Arseni et al., 2022). Also, cryo-EM images reveal the presence of both single and double TMEM106B filaments in each individual.
However, the presence of both TDP-43 filaments and TMEM106B filaments in type A FTLD-TDP and type B FTLD-TDP (Schweighauser et al., 2022) contradicts a recent report (Jiang et al., 2022) which showed that amyloid filaments in FTLD-TDP are composed of TMEM106B along with abundant non-fibrillar aggregated TDP-43, but not TDP-43 fibrils. The authors of the current report argue that differences in brain extraction protocols could explain this discrepancy. They found TDP43 filaments in the supernatant after centrifugation at 27,000 g, while the other study examined the pellet after centrifugation at 21,000 g. This is analogous to the discovery of abundant Aβ fibrils in ultracentrifugal supernatants of aqueous extracts from Alzheimer's disease brains (Stern et al., 2023).
The question then arises whether "purified" small aggregates or "protofibrils" from globular co-aggregates with other proteins inhibiting fibrillation can indeed form fibrils in the supernatant fraction. TDP may exhibit two distinct behaviors: N-terminus intact TDP has a high propensity to form functional oligomers-dimers that bind nucleic acids through RNA binding motifs, yet can misfold to generate globular aggregates, or co-aggregates, found in the pellets. In contrast, the amyloidogenic C-terminal (N-truncated) TDP forms “abundant” filaments found in the supernatant.
Another significant point of the current study is that individual filaments exhibit heterogeneity, and uniform repetitive structures are not always assembled. Various local structural variations could coexist within individual filaments. Importantly, this study and that of Kumar et al., 2023, highlights the role of post-translational modifications (PTMs) in shaping the local structural variation of TDP-43 filaments. These modifications determine the preferred structural form of the filament structure, indicating that PTMs occurring prior to fibrillization, and influenced by different neurodegenerative conditions, ultimately dictate the final fold of the filaments.
This concept is also applicable to other intrinsically disordered proteins, such as Aβ and tau. Thus, this suggests a broader concept where specific, disease-related PTM patterns determine the amyloid filament conformation of distinct proteins in neurodegenerative diseases.
References:
Arseni D, Hasegawa M, Murzin AG, Kametani F, Arai M, Yoshida M, Ryskeldi-Falcon B. Structure of pathological TDP-43 filaments from ALS with FTLD. Nature. 2022 Jan;601(7891):139-143. Epub 2021 Dec 8 PubMed.
Jiang YX, Cao Q, Sawaya MR, Abskharon R, Ge P, DeTure M, Dickson DW, Fu JY, Ogorzalek Loo RR, Loo JA, Eisenberg DS. Amyloid fibrils in FTLD-TDP are composed of TMEM106B and not TDP-43. Nature. 2022 May;605(7909):304-309. Epub 2022 Mar 28 PubMed.
Kumar ST, Nazarov S, Porta S, Maharjan N, Cendrowska U, Kabani M, Finamore F, Xu Y, Lee VM, Lashuel HA. Seeding the aggregation of TDP-43 requires post-fibrillization proteolytic cleavage. Nat Neurosci. 2023 Jun;26(6):983-996. Epub 2023 May 29 PubMed.
Schweighauser M, Arseni D, Bacioglu M, Huang M, Lövestam S, Shi Y, Yang Y, Zhang W, Kotecha A, Garringer HJ, Vidal R, Hallinan GI, Newell KL, Tarutani A, Murayama S, Miyazaki M, Saito Y, Yoshida M, Hasegawa K, Lashley T, Revesz T, Kovacs GG, van Swieten J, Takao M, Hasegawa M, Ghetti B, Spillantini MG, Ryskeldi-Falcon B, Murzin AG, Goedert M, Scheres SH. Age-dependent formation of TMEM106B amyloid filaments in human brains. Nature. 2022 May;605(7909):310-314. Epub 2022 Mar 28 PubMed.
Stern AM, Yang Y, Jin S, Yamashita K, Meunier AL, Liu W, Cai Y, Ericsson M, Liu L, Goedert M, Scheres SH, Selkoe DJ. Abundant Aβ fibrils in ultracentrifugal supernatants of aqueous extracts from Alzheimer's disease brains. Neuron. 2023 Jul 5;111(13):2012-2020.e4. Epub 2023 May 10 PubMed.
Biomedizinisches Centrum (BMC), Biochemie & Deutsches Zentrum für Neurodegenerative Erkrankungen (DZNE)
This manuscript by the Ryskeldi-Falcon lab clarifies several important issues. It is clear now that TDP-43 forms amyloid fibers in FTLD Type A and Type B—and, as in other neurodegenerative diseases, these fibers show distinct structures and are therefore disease defining. The unambiguous presence of TDP-43 filaments in both FTLD-TDP type A and B further supports the pathological nature of these aggregates. Most importantly, the findings in this paper, as well as in the earlier publication (Arseni et al., 2022), provide a basis for the development of disease-specific diagnostics and therapeutics.
Importantly, these findings are in contrast to previous observations that amyloid fibers in FTLD-TDP were composed solely of TMEM106B, but not of TDP-43 (Jiang et al., 2022).
But what about a pathological contribution of the TMEM106B deposits, if any? Their ubiquitous detection in many different neurodegenerative disorders, and their age-dependent occurrence even in neurologically normal humans, suggest that they are either innocent bystanders or that they enhance the risk for many different neurodegenerative diseases by an unknown mechanism.
The rather ubiquitous occurrence of the TMEM106B deposits in many neurological conditions would suggest a disease-overarching mechanism. Based on the age-dependent occurrence of the TMEM106B deposits, I could imagine that they may contribute to disease risk by affecting age-related processes in the brain and therefore facilitate earlier disease onset. One possibility is that the TMEM106B fibers arise in dysfunctional endosomes/lysosomes and may then even further reduce lysosomal function. Over time, this may then lead to disruption of lysosomes and leakage of lysosomal content into the cytoplasm.
References:
Arseni D, Hasegawa M, Murzin AG, Kametani F, Arai M, Yoshida M, Ryskeldi-Falcon B. Structure of pathological TDP-43 filaments from ALS with FTLD. Nature. 2022 Jan;601(7891):139-143. Epub 2021 Dec 8 PubMed.
Jiang YX, Cao Q, Sawaya MR, Abskharon R, Ge P, DeTure M, Dickson DW, Fu JY, Ogorzalek Loo RR, Loo JA, Eisenberg DS. Amyloid fibrils in FTLD-TDP are composed of TMEM106B and not TDP-43. Nature. 2022 May;605(7909):304-309. Epub 2022 Mar 28 PubMed.
The University of Sydney
In this important analysis, Arseni et al. build on their previous work. They demonstrate that the fibrils underpinning the two most common TDP-43 subtypes are composed of distinct folds of the same LCD region—chevron-like in type A and spiral-like in type B—of which the former may have been facilitated by post-translational modifications of arginine 293.
These findings provide structural support for the pathological distinction of FTLD-TDP types A and B, which are often the most difficult to differentiate, even by the most experienced neuropathologists (Alafuzoff et al., 2015). It would be interesting to know whether the filament folds of TDP-43 in ALS are indeed the same as those described in FTLD-ALS TDP type B, and if this differs from type E, which has a more distinctive morphology, distribution, and spread compared to types A and B (Porta et al., 2021), and which we have found more commonly represented in ALS cases without FTLD.
University of Florence
This manuscript consolidates previous findings by the same group that TDP-43 can form filaments with β-strands that stack in a parallel, in-register arrangement (chevron fold), although with a different fold with respect to the previously described filaments (double-spiral-shaped fold). It removes doubts that previously found filaments reported by the same group last year (Arseni et al., 2022) derived from the TMEM106B protein.
Benjamin Ryskeldi-Falcon, whom I had the pleasure of contacting last year to discuss this specific point, was well aware of the presence of TMEM106b fibrils in the elderly and ruled out, after his first Nature paper was released in 2022, that his TDP-43 filaments consisted of TMEM106B fibrils.
A doubt remains that many of the TDP-43 inclusions may not have filaments altogether, particularly the round inclusions, as opposed to the skein-like inclusions. Robinson et al. in 2013 had noticed that only a limited subset of TDP-43 skein-like inclusions could bind weakly Thioflavin S, whereas none of the round inclusions did. Papers by Jiang et al. and Laferrière et al. in Nature and Nature Neuroscience in 2022 and 2016, respectively, showed absence of TDP-43 filaments. I saw comments that this may arise from the different extraction procedures, but I wonder whether this is due to the polymorphism in TDP-43 aggregation that lead to non-filamentous structure in many of the cases, as well as different folds when filaments do form.
We indeed had trouble generating filamentous structures by expressing TDP-43 in cells or from the purified protein, indicating its low propensity to form amyloid filaments (Cascella et al., 2023).
It's also important to discuss that neither of the TDP-43 filament folds described in 2022 and 2023 possess a clear stacking of β-sheets along an axis perpendicular to the major filament axis. In X-ray fiber diffraction of oriented fibrils, the TDP-43 filaments would show the 4.7 Å reflection on the meridional axis, but not the 10-11 Å reflection on the equatorial axis, which has been used in the past to classify aggregated fibrils as amyloid. Absence of thioflavin S/T binding also represents a departure from classical fibrils.
References:
Arseni D, Hasegawa M, Murzin AG, Kametani F, Arai M, Yoshida M, Ryskeldi-Falcon B. Structure of pathological TDP-43 filaments from ALS with FTLD. Nature. 2022 Jan;601(7891):139-143. Epub 2021 Dec 8 PubMed.
Jiang YX, Cao Q, Sawaya MR, Abskharon R, Ge P, DeTure M, Dickson DW, Fu JY, Ogorzalek Loo RR, Loo JA, Eisenberg DS. Amyloid fibrils in FTLD-TDP are composed of TMEM106B and not TDP-43. Nature. 2022 May;605(7909):304-309. Epub 2022 Mar 28 PubMed.
Laferrière F, Maniecka Z, Pérez-Berlanga M, Hruska-Plochan M, Gilhespy L, Hock EM, Wagner U, Afroz T, Boersema PJ, Barmettler G, Foti SC, Asi YT, Isaacs AM, Al-Amoudi A, Lewis A, Stahlberg H, Ravits J, De Giorgi F, Ichas F, Bezard E, Picotti P, Lashley T, Polymenidou M. TDP-43 extracted from frontotemporal lobar degeneration subject brains displays distinct aggregate assemblies and neurotoxic effects reflecting disease progression rates. Nat Neurosci. 2019 Jan;22(1):65-77. Epub 2018 Dec 17 PubMed.
Robinson JL, Geser F, Stieber A, Umoh M, Kwong LK, Van Deerlin VM, Lee VM, Trojanowski JQ. TDP-43 skeins show properties of amyloid in a subset of ALS cases. Acta Neuropathol. 2013 Jan;125(1):121-31. PubMed.
Cascella R, Banchelli M, Abolghasem Ghadami S, Ami D, Gagliani MC, Bigi A, Staderini T, Tampellini D, Cortese K, Cecchi C, Natalello A, Adibi H, Matteini P, Chiti F. An in situ and in vitro investigation of cytoplasmic TDP-43 inclusions reveals the absence of a clear amyloid signature. Ann Med. 2023 Dec;55(1):72-88. PubMed.
Make a Comment
To make a comment you must login or register.