Could Untranslated Regions of RNA Help Control Gene Expression?
Quick Links
Scientists may have uncovered a new way for cells to regulate protein expression. In the December 16 Neuron, researchers led by Mary Hynes at Rockefeller University, New York, reported that in mouse neurons, the 3' untranslated regions of mRNAs were often expressed at higher or lower levels than the corresponding coding regions of the same strand. These expression patterns stayed consistent within particular neuronal subtypes and between animals, but varied with age and brain region, suggesting they were not random and might serve a purpose. Hinting at this, preliminary data on two genes linked high expression of the 3'UTRs to low expression of the encoded proteins. If the results hold up for more genes, this would add yet another wrinkle for geneticists trying to analyze gene expression.
Other researchers called the findings intriguing but preliminary. “This is a very interesting study which may give rise to new ways of thinking about the relationship of mRNA sequence to protein expression,” Bruce Yankner at Harvard Medical School wrote to Alzforum. Likewise, Philip De Jager at Brigham and Women’s Hospital, Boston, noted, “Often, the correlation between mRNA and protein levels is not very strong. This may explain part of the discrepancy we see.” However, both commenters noted that the sample size was limited, and suggested that researchers will need to look at many more genes in a systematic manner to determine if this represents a widespread regulatory phenomenon.
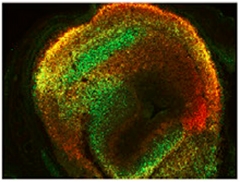
Patterns of Unequal Expression.
In mouse forebrain, researchers saw distinct sets of cells where Sox11 mRNA-untranslated regions were highly expressed (red), minimally expressed (green), or expressed in equal measure with coding regions (yellow). [Courtesy of Neuron, Kocabas et al.]
Hynes and colleagues first noticed the phenomenon serendipitously, while using translating ribosome affinity purification (TRAP) mice to study gene expression in developing dopamine neurons. These mice, developed by Nathaniel Heintz and colleagues at Rockefeller, allow researchers to selectively isolate mRNA from specific classes of neurons that express a particular promoter (see Heiman et al., 2008; Heiman et al., 2014). They do this by using a ribosome tag, such as enhanced green fluorescent protein/ribosomal subunit chimera, driven by that promoter. Researchers have used TRAP to examine RNA expression patterns from distinct subtypes of neurons, without having to isolate them first. This technology made the unequal mRNA expression patterns visible, Hynes told Alzforum.
Joint first authors Arif Kocabas and Terence Duarte used dopamine transporter promoter-based TRAP mice to isolate mRNA from mouse embryonic dopamine neurons. When they sequenced the mRNA, they noticed that for many transcripts there were thousands of copies of the 3'UTR, but virtually none of the coding region. To see if this was a common occurrence, the authors performed in situ hybridizations using probes to the coding and 3'UTR sequences of 19 genes, examining tissue from multiple brain regions and peripheral organs of mice, and from animals of different ages. The genes ranged from highly expressed housekeeping genes to transiently expressed developmental genes. In every case, the authors found unequal expression of coding regions and 3'UTRs at some ages and in some tissues. For each gene, the pattern stayed consistent across animals. In many cases, the authors saw precise spatial regulation, with adjacent subclasses of neurons possessing distinct expression patterns (see image above).
What purpose might these patterns serve? The authors compared mRNA to protein levels for two genes, tyrosine hydroxylase and microtubule-associated protein 2, for which they had good antibodies. For both genes, the authors found that the expression level of the protein product in specific tissues was not correlated with total mRNA levels, but instead with the ratio of coding sequence to 3'UTR. Cells with high levels of 3'UTRs expressed little to none of the corresponding protein product. The results hint that an excess of untranslated mRNA could help turn down unneeded protein expression, the authors suggest. Supporting this, an analysis of RNA sequencing data from the embryonic dopaminergic neurons revealed that the set of genes with high 3'UTR to coding region ratios was enriched for developmental genes, whose precise regulation is critical to development of various cells and organs.
Exactly how a cell acquires an excess of 3'UTR without its corresponding coding region remains unclear. A previous study by researchers led by John Mattick at the University of Queensland, Brisbane, Australia, found that 3'UTRs can be expressed separately from coding regions through post-transcriptional cleavage and degradation of the coding portions of the mRNA (see Mercer et al., 2011). These UTRs might function as noncoding RNA and regulate gene expression, the Australian group speculated. Hynes plans to further investigate this potential function by overexpressing 3'UTRs in cultured cells, and measuring whether this modulates protein expression. She will also examine more genes to determine if overexpression of UTRs consistently correlates with silencing.—Madolyn Bowman Rogers
References
Paper Citations
- Heiman M, Schaefer A, Gong S, Peterson JD, Day M, Ramsey KE, Suárez-Fariñas M, Schwarz C, Stephan DA, Surmeier DJ, Greengard P, Heintz N. A translational profiling approach for the molecular characterization of CNS cell types. Cell. 2008 Nov 14;135(4):738-48. PubMed.
- Heiman M, Kulicke R, Fenster RJ, Greengard P, Heintz N. Cell type-specific mRNA purification by translating ribosome affinity purification (TRAP). Nat Protoc. 2014;9(6):1282-91. Epub 2014 May 8 PubMed.
- Mercer TR, Wilhelm D, Dinger ME, Soldà G, Korbie DJ, Glazov EA, Truong V, Schwenke M, Simons C, Matthaei KI, Saint R, Koopman P, Mattick JS. Expression of distinct RNAs from 3' untranslated regions. Nucleic Acids Res. 2011 Mar;39(6):2393-403. Epub 2010 Nov 12 PubMed.
Further Reading
News
- ENCODE Expands Analysis of DNA Regulation
- Listen Up, Gene Silencing Strikes a Chord at RNA Meeting
- Amplitude Modulation—Do MicroRNAs Muffle Noise in Gene Expression?
- Do MicroRNAs Cause Mayhem Across Frontotemporal Dementia Spectrum?
- Glymphatic Flow, Sleep, microRNA Are Frontiers in Alzheimer’s Research
Primary Papers
- Kocabas A, Duarte T, Kumar S, Hynes MA. Widespread Differential Expression of Coding Region and 3' UTR Sequences in Neurons and Other Tissues. Neuron. 2015 Dec 16;88(6):1149-56. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Kentucky
University of Kentucky
The study by Kocabas et al., revealing “Widespread differential expression of coding region (CDS) and 3'UTR sequences in neurons ...” for many genes is both surprising and highly interesting. According to current thinking, the 3'UTR and CDS of a gene are expected to show relatively similar abundance, as they are different regions of the same mRNA. However, the authors present convincing in situ hybridization and other evidence that this expectation in many cases may well be incorrect. The study reveals several surprising results, including: 1) that there are different levels of 3'UTR to CDS for some, but not all genes; 2) that these occur in patterns that are developmentally and spatially regulated and conserved across different biological samples; and 3) that there is a correlation between the 3'UTR-to-CDS ratio and protein level. This phenomenon is widespread among genes, including several genes that are commonly used as “control” genes (β-actin and GAPDH). But the unbalance is not random, in that certain gene ontology categories are more likely to be associated with the 3'UTR disconnect than others.
The functional implications of these new findings are not yet clear. The data could indicate that post-translational degradation or pre-translational association with ribosomes of some mRNA species involves previously unrecognized cleavage of the UTR and CDS. Alternatively, the correlation between protein and UTR-to-CDS ratio may indicate that this unbalance reflects a novel mechanism of gene regulation, perhaps one designed to coordinate rapid alterations in expression of related genes by separating the CDS from the regulatory influence of the 3'UTR in multiple sensitive genes of a functional network. If further work supports the conclusion that this differential expression reflects regulatory mechanisms, then, as the authors note, probe sets for all regions of the mRNA will need to be included in microarrays. And then it will be of great interest to determine which gene categories and networks are subject to such regulation and whether neurons vulnerable to neurodegenerative disorders are particularly susceptible (or resistant) to this type of differential expression.
Nathan Kline Institute/NYU Langone School of Medicine
This report of differential expression of exon-coding sequences (CDS) and 3′ UTR sequences in neurons and other cellular sources provides interesting information regarding potential regulation of cell-specific expression of individual transcripts and their encoded proteins during development and in adulthood. The work has strong implications for molecular mechanisms underlying neurodegenerative disorders. Interestingly, defined neuronal populations were found to have varying ratios of cognate 3′ UTR and CDS sequences, as some genes displayed a high 3′ UTR:CDS ratio while others revealed a low 3′ UTR:CDS ratio. A functional consequence of a differential 3′ UTR:CDS ratio was demonstrated in two genes whereby a high ratio correlated strongly with low protein expression. The discrepant levels of cognate 3′ UTR and CDS sequences is somewhat surprising, as the generally accepted dogma is that these elements were more or less equivalent until translation or degradation. The authors are commended for looking at neurons in different brain regions during development and adulthood as well as other tissue sources, such as the kidney.
Discrete expression of tyrosine hydroxylase (TH) 3′ UTR:CDS ratio and protein in embryonic midbrain neurons provided convincing proof of concept that a ratio >3 predicted low or no protein expression. Extrapolating this finding of cell-specific 3′ UTR:CDS ratios to AD pathogenesis, it will be interesting to evaluate cognate 3′ UTR:CDS sequences in neurons that are selectively vulnerable to neurodegeneration during the progression of AD. For example, cholinergic markers such as choline acetyltransferase and neurotrophin receptors such as TrkA and p75NTR in cholinergic basal forebrain (CBF) neurons in AD, mild cognitive impairment (MCI), and age-matched nondemented controls would be ideal to evaluate potential pathogenic shifts in 3′ UTR:CDS ratios. Parallel studies would be of high interest in animal models of AD pathology where basal forebrain cholinergic neurons (BFCNs) lose their phenotype, including the Ts65Dn mouse model of AD and Down’s syndrome.
In sum, the data present by Kocabas et al. has broad relevance for AD research. Re-evaluation of high-throughput (e.g., RNA-seq and/or microarray) data as well as prospective studies with built-in cognate 3′ UTR and CDS measurements is warranted to parse out potential large-scale expression discrepancies between 3′ UTR and CDS sequences. A caveat would be that data must arise from individual populations, e.g., from neurons extracted via laser-capture microdissection rather than regional or global tissue dissections to avoid contaminating signals from different neuronal, glial, and vascular cell populations, among others. Unfortunately, calculating the 3′ UTR:CDS ratio from data repositories using admixed cell types in postmortem AD tissues or AD animal models will not likely lead to interpretable results based upon the analyses of Kocabas et al., because the 3′ UTR:CDS ratio is likely cell-type specific throughout development and adulthood.
Make a Comment
To make a comment you must login or register.