Acetylation Accelerates Aggregation of 3R, but Not 4R, Tau
Quick Links
Tauopathies are classified by which tau isoform forms fibrils—those with three microtubule-binding domain repeats (3R), those with four (4R), or those with both. What drives one isoform to aggregate over the other? Post-translational modifications, namely acetylation, say researchers led by Markus Zweckstetter of the German Center for Neurodegenerative Diseases in Göttingen. In the September 22 Nature Communications, they reported that acetylation drives 3R tau to form fibrils in vitro while it prevents the 4R isoform from aggregating. Why? Acetylated lysine residues within a segment unique to 4R tau disrupted a fold that stabilizes fibrils. Zweckstetter thinks this modification might explain why only 3R tau deposits in the brains of people with Pick’s disease, a 3R tauopathy.
“This paper presents a plausible mechanism for selective 3R tau deposition,” wrote Marc Diamond, University of Texas Southwestern Medical Center, Dallas. “However, before we can understand this, we would need to study the process in cells.”
Lukasz Joachimiak, also at the University of Texas in Dallas, agreed. “I think it’s too early to say how post-translational modifications interface with tau fibril formation and link it to disease,” he wrote.
Thirty-five percent of tau’s residues can be adorned with acetate, phosphate, and other post-translational modifications (reviewed by Alquezar et al., 2020). Scientists have spotted distinct acetylation patterns in brain tissue from people with different tauopathies, including Pick’s and Alzheimer’s diseases (image below; Feb 2020 news; Kametani et al., 2020). It is unclear how those patterns relate to the uniquely shaped tau fibrils that form in different tauopathies.
To learn more about tau acetylation and how it affects aggregation, first author Pijush Chakraborty acetylated recombinant 3R and 4R tau by adding the lysine acetyltransferases p300 and CBP to tau in solution. Chakraborty chose these enzymes because they are dysregulated in tauopathies. In the AD brain, they are downregulated (Schueller et al., 2020). Three-R tau comprises microtubule-binding domains 1, 3, and 4, while 4R tau contains the second repeat as well.
Twelve hours later, almost all the 37 3R and 44 4R lysines were modified, with K317 and K321 being the most strongly acetylated in both isoforms. Notably, all five lysine residues in repeat 2 were acetylated. Three of these residues—K290, K294, and K298—were also acetylated when tau mingled with the acetylases in the presence of microtubules, suggesting these modifications might take place in vivo.
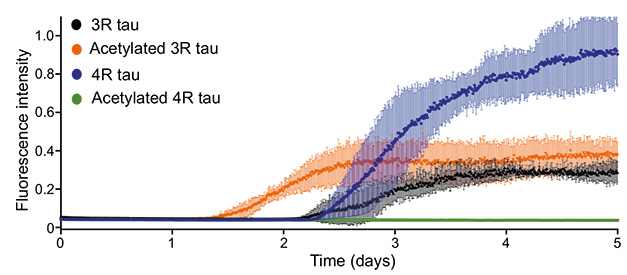
Slow, slow, quick. Compared to unadorned 3R tau (black), the acetylated version (orange) fibrillized faster. Four-R tau (blue) aggregates more than either form of 3R tau, but acetylated 4R tau (green) does not aggregate at all. [Courtesy of Chakraborty et al., Nature Communications, 2023.]
How did acetylation affect tau aggregation? Naked 3R and 4R tau began aggregating, as measured by thioflavin T (ThT) fluorescence, after two days. Four-R tau formed three times as many fibrils as did 3R tau after five days. Acetylation accelerated 3R tau aggregation, with ThT fluorescence being detectable after 1.5 days (image above). The scientists think that, by capping the positively charged lysines, acetylation removes electrostatic repulsion, driving 3R fibril formation. Joachimiak agreed. “The modification probably contributes to nucleation by stabilizing a conformation that is aggregation-prone or destabilizing one resistant to aggregation,” he wrote. Joachimiak does not think acetylation accelerates the elongation of fibrils because the amount of acetylated tau in these structures is low.
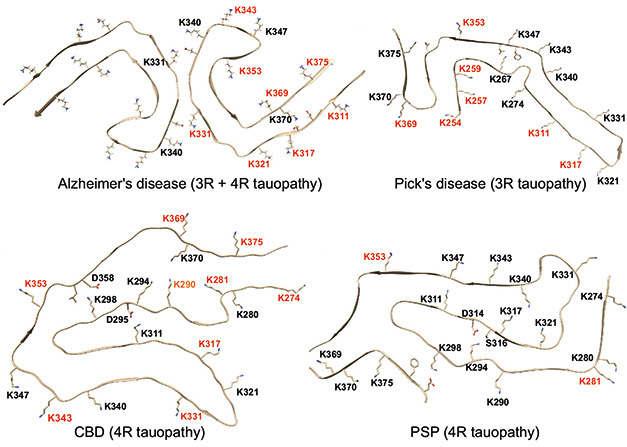
Acetylation Across Tauopathies. Different lysine residues are acetylated (red) among various tau protofibril folds. [Courtesy of Chakraborty et al., Nature Communications, 2023.]
Surprisingly, acetylated 4R tau formed no aggregates. Why? The authors think acetylation in the second microtubule-binding repeat must explain this observation, since it is unique to the 4R isoform. To test how each of these lysines individually affects aggregation, they mutated each in turn to cysteine, then chemically converted them to N-acetylcysteamine, which mimics acetylated lysine. Each of the four mimics individually delayed tau aggregation, with the K298 variant stalling fibril formation the longest, until about four days.
In cryoEM-determined structures of tau fibrils from people with corticobasal degeneration (CBD), this positively charged lysine reaches in, toward the core of the protofibril fold, to hold electrostatic hands with a negatively charged aspartic acid, stabilizing the fibril (image above). To the authors, this explains why acetylation slows 4R aggregation and why it selectively drives 3R tauopathies, such as Pick’s disease.—Chelsea Weidman Burke
References
News Citations
Paper Citations
- Alquezar C, Arya S, Kao AW. Tau Post-translational Modifications: Dynamic Transformers of Tau Function, Degradation, and Aggregation. Front Neurol. 2020;11:595532. Epub 2021 Jan 7 PubMed.
- Kametani F, Yoshida M, Matsubara T, Murayama S, Saito Y, Kawakami I, Onaya M, Tanaka H, Kakita A, Robinson AC, Mann DM, Hasegawa M. Comparison of Common and Disease-Specific Post-translational Modifications of Pathological Tau Associated With a Wide Range of Tauopathies. Front Neurosci. 2020;14:581936. Epub 2020 Nov 4 PubMed.
- Schueller E, Paiva I, Blanc F, Wang XL, Cassel JC, Boutillier AL, Bousiges O. Dysregulation of histone acetylation pathways in hippocampus and frontal cortex of Alzheimer's disease patients. Eur Neuropsychopharmacol. 2020 Apr;33:101-116. Epub 2020 Feb 11 PubMed.
Further Reading
Primary Papers
- Chakraborty P, Rivière G, Hebestreit A, de Opakua AI, Vorberg IM, Andreas LB, Zweckstetter M. Acetylation discriminates disease-specific tau deposition. Nat Commun. 2023 Sep 22;14(1):5919. PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
University of Texas, Southwestern Medical Center
I think this very interesting paper presents a plausible mechanism for selective 3R deposition; however, before we can really understand this, we would need to study the process in cells. Also, the spontaneous aggregation of full-length tau in solution is unlikely to reproduce the folding patterns that occur in the brain, and this was not actually tested here.
So, within the limits of the study, I think this work suggests how a post-translational modification could change the folding of one form of tau versus another. A lot more work will be needed to try to extrapolate these findings to humans.
Mayo Clinic
These intriguing results provide compelling insight into the differential effect of acetylation on assembly of 3R and 4R tau isoforms. The inhibitory effect of acetylation on aggregation of 4R tau is consistent with our previous studies (Cook et al., 2014; Carlomagno et al., 2017), although we neglected to examine the impact of acetylation on assembly kinetics of 3R tau.
The authors’ discovery that acetylation drives aggregation of 3R yet inhibits aggregation of 4R offers a potential mechanism for the sole 3R tauopathy, Pick’s disease. In future studies, it would be interesting to assess acetylation of 3R and 4R tau isoforms in soluble and insoluble fractions from patients with Pick’s disease. The results described by Chakraborty and associates would predict that increased acetylation of soluble 4R tau and insoluble 3R would be observed in affected brain regions of people with this illness.
In addition, following the authors’ discovery of K298 as a hot spot for isoform-specific aggregation, the development of an acetylated-K298 site-specific antibody could be useful to better define the disease relevance of this modification. Finally, along with monitoring differential effects of acetylation on tau assembly, in future studies it would be interesting to examine the effect(s) of acetylation on tau function in an isoform-specific manner.
References:
Carlomagno Y, Chung DC, Yue M, Castanedes-Casey M, Madden BJ, Dunmore J, Tong J, DeTure M, Dickson DW, Petrucelli L, Cook C. An acetylation-phosphorylation switch that regulates tau aggregation propensity and function. J Biol Chem. 2017 Sep 15;292(37):15277-15286. Epub 2017 Jul 31 PubMed.
Cook C, Carlomagno Y, Gendron TF, Dunmore J, Scheffel K, Stetler C, Davis M, Dickson D, Jarpe M, Deture M, Petrucelli L. Acetylation of the KXGS motifs in tau is a critical determinant in modulation of tau aggregation and clearance. Hum Mol Genet. 2013 Aug 26; PubMed.
UT Southwestern Medical Center
It has been pretty tough to assign clear contributions of specific PTMs on tau in the disease process. Phosphorylation is relatively abundant on tau in pathological states, but is very heterogeneous. By contrast, acetylation is low abundance (but certainly enriched) in disease states compared to normal states.
If this is true, it’s probably not linked to end-stage aggregates, but rather contributes to nucleation where the modification stabilizes a conformation that is aggregation-prone (or conversely destabilizes an aggregation-resistant conformation) that is on path to a specific structural polymorph.
We recently published a paper in Structure identifying patterns of acetylation that drive tau fibril formation that were consistent with acetylation site patterns identified in AD, but their abundance in disease is low. We determined a cryo-EM structure of a fibril, but ultimately it’s not clear what this PTM pattern means for disease initiation and progression as the disease states do not have this modification on every single monomer in the fibril. This has been a challenge.
There are new ways that enable introduction of PTMS using orthogonal incorporation of amino acids at amber codons, but these methods so far cannot incorporate many substitutions at a time. I think it’s too early to say how PTMs interface with tau fibril formation and its link to disease.
Weill Cornell Medicine
These findings are very intriguing. Given the importance of 3R and 4R ratios in neurodegeneration, the potential differential effects of acetylation on 3R and 4R could have important implications and warrant further exploration in cell-based studies and in vivo. For example, in cell-based studies, expression of p300 and CBP greatly enhanced 4R tau acetylation, but did not lead to tau aggregation. It would be worthwhile to perform direct comparison with 3R tau and assess how tau aggregation is affected.
While it has not been shown to enhance tau aggregation, per se, acetylation of 4R tau could exert detrimental effects. We and others have used pseudo-acetylation 4R tau constructs, and we observed detrimental effects of aberrant acetylation of ac-K274, which is found in 3R and 4R isoforms, and ac-K281, which is unique to 4R tau (Tracy et al., 2016; Yan et al., 2022). It would be interesting to compare the effects of ac-K274 in 3R tau versus 4R tau, for example.
It is worth noting that 4R aggregates are enriched with ac-tau species in human tauopathy brains, including 4R tauopathies such as PSP and AD, a 3R/4R tauopathy. For example, an Ac-K280 antibody (4R-specific epitope) has detected tau aggregates in AD and other tauopathies (Irwin et al. 2012; Irwin et al., 2013). Acetylated-tau (ac-K274, 3R/4R shared epitope) is enriched in tau aggregates in both PSP, as well as the 3R tauopathy, Pick’s disease (Grinberg et al., 2013). A more recent quantitative MS study further expanded and confirmed the enrichment of tau acetylation in AD, including epitopes unique to 4R (Wesseling et al., 2020).
References:
Tracy TE, Sohn PD, Minami SS, Wang C, Min SW, Li Y, Zhou Y, Le D, Lo I, Ponnusamy R, Cong X, Schilling B, Ellerby LM, Huganir RL, Gan L. Acetylated Tau Obstructs KIBRA-Mediated Signaling in Synaptic Plasticity and Promotes Tauopathy-Related Memory Loss. Neuron. 2016 Apr 20;90(2):245-60. Epub 2016 Mar 31 PubMed.
Yan Y, Wang X, Chaput D, Shin MK, Koh Y, Gan L, Pieper AA, Woo JA, Kang DE. X-linked ubiquitin-specific peptidase 11 increases tauopathy vulnerability in women. Cell. 2022 Oct 13;185(21):3913-3930.e19. Epub 2022 Oct 4 PubMed.
Irwin DJ, Cohen TJ, Grossman M, Arnold SE, Xie SX, Lee VM, Trojanowski JQ. Acetylated tau, a novel pathological signature in Alzheimer's disease and other tauopathies. Brain. 2012 Mar;135(Pt 3):807-18. PubMed.
Irwin DJ, Cohen TJ, Grossman M, Arnold SE, McCarty-Wood E, Van Deerlin VM, Lee VM, Trojanowski JQ. Acetylated tau neuropathology in sporadic and hereditary tauopathies. Am J Pathol. 2013 Aug;183(2):344-51. PubMed.
Grinberg LT, Wang X, Wang C, Sohn PD, Theofilas P, Sidhu M, Arevalo JB, Heinsen H, Huang EJ, Rosen H, Miller BL, Gan L, Seeley WW. Argyrophilic grain disease differs from other tauopathies by lacking tau acetylation. Acta Neuropathol. 2013 Apr;125(4):581-93. PubMed.
Wesseling H, Mair W, Kumar M, Schlaffner CN, Tang S, Beerepoot P, Fatou B, Guise AJ, Cheng L, Takeda S, Muntel J, Rotunno MS, Dujardin S, Davies P, Kosik KS, Miller BL, Berretta S, Hedreen JC, Grinberg LT, Seeley WW, Hyman BT, Steen H, Steen JA. Tau PTM Profiles Identify Patient Heterogeneity and Stages of Alzheimer's Disease. Cell. 2020 Dec 10;183(6):1699-1713.e13. Epub 2020 Nov 13 PubMed.
Make a Comment
To make a comment you must login or register.