San Francisco: Is Tau Reduction a Good Thing?
Quick Links
Though amyloid-β and tau stand as the pathological hallmarks of Alzheimer’s disease, only recently have scientists gained insight into how the two could be linked mechanistically. Research from several labs suggests that tau acts downstream of Aβ. What’s more, recent studies have shown that animals lacking tau are impervious to the synaptic and cognitive problems normally brought on by Aβ overproduction, raising the notion of tau reduction as a potential therapeutic avenue. At the “Tau and Tauopathies: Pathogenic Mechanisms” workshop held 28-30 March 2011 at the Gladstone Institute for Neurological Disease in San Francisco (see Part 1), several speakers presented new data in this vein. Some of it complicates the picture, suggesting that tau reduction may not always be beneficial.
Early hints of tau’s essential role in Aβ’s dirty work came from cell culture studies showing that tau-depleted hippocampal neurons do not wither despite the presence of Aβ (Rapoport et al., 2002). Erik Roberson, while a postdoc in Lennart Mucke’s lab at the Gladstone Institute, confirmed those findings in vivo by showing that halving tau levels in APP-overexpressing J20 mice kept them from dying prematurely and helped them resist the Aβ-induced cognitive impairment (ARF related news story on Roberson et al., 2007). In addition, these scientists and others report that reducing tau protects APP mice against axonal transport defects (ARF related news story on Vossel et al., 2010) relieves their spontaneous epileptiform activity (ARF related news story on Roberson et al., 2011) and lessens Aβ-induced impairment of hippocampal long-term potentiation (Shipton et al., 2011). The tyrosine kinase Fyn may contribute to these effects, as a recent study finds that tau targets Fyn to post-synaptic N-methyl-D-aspartic acid (NMDA) receptors (ARF related news story on Ittner et al., 2010; see also review by Ittner and Götz, 2011).
At the meeting, Roberson, who now heads a lab at the University of Alabama at Birmingham, presented new data suggesting that the benefits of tau reduction may extend to APP mice that express a Fyn transgene but have low Aβ levels. Despite their low Aβ production, the APP/Fyn mice develop cognitive impairment, and almost half die within six months. However, putting these mice onto a tau knockout background rescued the mortality phenotype, Roberson said.
More evidence that tau reduction could be beneficial apart from Aβ came from Yadong Huang of the Gladstone Institute. When his group analyzed apolipoprotein E knock-in mice expressing either the human E3 or E4 isoform, the scientists found that E4 knock-in mice selectively lost GABAergic interneurons to an extent that correlated with the animals’ learning and memory deficits (Andrews-Zwilling et al., 2010). The GABAergic interneurons that did remain in the E4 mice had excessive tau phosphorylation, and the scientists were able to enhance their survival and prevent cognitive impairment by knocking down endogenous tau with microRNAs.
Tau also appears necessary for a pyroglutamate form of Aβ (pyroGluAβ) to inflict its harm on neurons. This is according to research by George Bloom, University of Virginia, Charlottesville, previously presented at the 2010 Society for Neuroscience meeting in San Diego, California. PyroGluAβ is a highly stable, neurotoxic species that arises when an enzyme joins the ends of a glutamate residue exposed by removal of Aβ42’s first two N-terminal amino acids. Bloom’s team, including former graduate student Justin Nussbaum and Probiodrug AG scientists led by Hans-Ulrich Demuth, showed that oligomers made by mixing Aβ42 with a touch of pyroGluAβ killed primary cortical neurons in culture more potently than either peptide by itself. However, these oligomers hardly posed a threat to cultured neurons from tau knockout mice, suggesting tau is needed for pyroGluAβ aggregates to become menacing (see ARF conference story). “Maybe amyloid is the trigger that gets pathology started,” Bloom said in San Francisco. “But once that trigger is pulled, tau must be present for Aβ to do its damage.”
Lest you think removing tau improves everything, data from Hana Dawson and coworkers at Duke University in Durham, North Carolina, gives reason for pause. When taken into culture, primary hippocampal neurons from tau-deficient mice mature with a delay (Dawson et al., 2001), and hippocampal neurons from tau-/- Map1b-/- mice do not extend their axons or migrate properly (Takei et al., 2000). Furthermore, crossing Tg2576 APP mice onto a tau knockout background gave rise to dystrophic neurites and axon degeneration—effects rescued by mating the APP strain to human tau transgenic mice. Traumatic brain injury (TBI) hastened the neuritic and axonal impairments, and the effects were intensified in APP mice on a tau knockout, relative to tau heterozygote background (Dawson et al., 2010) (see image below). This pertains to the emerging field of chronic traumatic encephalopathy, a tauopathy in concussed athletes and military veterans with TBI, Dawson noted in an e-mail to ARF (see also ARF Live Discussion and ARF related news story on TBI).
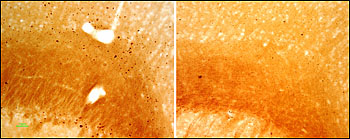
Among Tg2576 APP mice receiving traumatic brain injury at six months of age, those on a tau-deficient background (left) had more clumps of misshapen axons (aka “spheroids”) than tau did heterozygote controls (right). Image credit: Hana Dawson
Dawson also reported preliminary data showing that loss of tau worsens motor defects and brings on disease earlier in an ALS mouse model that overexpresses mutant SOD1 (A93G). To complicate matters, an earlier study found that tau deficiency did not matter much, good or bad, for the mouse central nervous system (Harada et al., 1994). “Removing tau can be a double-edged sword,” Dawson said. In discussion, scientists noted that the impact of tau reduction remains to be fully established. Some wondered whether strain differences may underlie the discrepancies between the various studies.
Dawson closed by noting that tau knockout mice probably have compensatory mechanisms. “How much worse would knockout of tau be in an adult?” she asked. This, as opposed to gene deficiency from birth, may more closely resemble a tau-reducing therapy started in people already afflicted with tauopathies.
Mucke hopes to address this issue using a new mouse model with regulatable neuronal expression of anti-tau microRNA that his group generated. Feeding the mice doxycycline keeps a lid on the microRNA, allowing endogenous tau to be expressed, but withdrawing doxycycline for just one week made an appreciable dent in tau expression. “So far, we see no obvious neurological deficits or other phenotypic changes,” Mucke reported. “Detailed analyses of these mice are in progress.” The data will be compared to those to come from the Roberson lab, which has generated conditional tau knockout mice using a Cre/Lox approach. In the meantime, Mucke and colleagues are screening RNA interference and drug libraries to identify tau-reducing reagents.
In a rare mention of “amyloid cascade hypothesis” at this tau meeting, Simon Lovestone of King’s College London, U.K., said “we have paid too much attention to ‘amyloid’ and not enough to ‘cascade.’” To address this shortcoming, Lovestone discussed how Dkk1—a Wnt signaling inhibitor that is increased in AD and in mouse models of neurodegenerative disease (Caricasole et al., 2004; Rosi et al., 2010)—may fit into this cascade. Lovestone and colleagues used whole-gene expression arrays to identify genes responsive to Dkk1, and to Aβ. The shared genes were induced in amyloidogenic transgenic mice (Tg2576) but not in a tauopathy model (hTau), Lovestone reported. This trend also held in their analyses of postmortem human brain tissue, supporting the idea that the shared gene set “sits neatly between amyloid and tau,” Lovestone said.—Esther Landhuis.
This is Part 3 of a four-part series. See also See also Part 1, Part 2, Part 4. Read the PDF of the entire series.
References
News Citations
- San Francisco: Gladstone Institute Hosts Tau Powwow
- APP Mice: Losing Tau Solves Their Memory Problems
- The Plot Thickens: The Complicated Relationship of Tau and Aβ
- Tau’s Synaptic Hats: Regulating Activity, Disrupting Communication
- Honolulu: The Missing Link? Tau Mediates Aβ Toxicity at Synapse
- San Diego: Pilin’ on the Pyro, Aβ Going Rogue
- Stress and Trauma: New Frontier Lures Alzheimer’s Researchers
- San Francisco: Making Tau Toxic—Post-translational Changes Galore
- San Francisco: Tau—Time to Shine as Therapeutic Target?
Webinar Citations
Paper Citations
- Rapoport M, Dawson HN, Binder LI, Vitek MP, Ferreira A. Tau is essential to beta -amyloid-induced neurotoxicity. Proc Natl Acad Sci U S A. 2002 Apr 30;99(9):6364-9. PubMed.
- Roberson ED, Scearce-Levie K, Palop JJ, Yan F, Cheng IH, Wu T, Gerstein H, Yu GQ, Mucke L. Reducing endogenous tau ameliorates amyloid beta-induced deficits in an Alzheimer's disease mouse model. Science. 2007 May 4;316(5825):750-4. PubMed.
- Vossel KA, Zhang K, Brodbeck J, Daub AC, Sharma P, Finkbeiner S, Cui B, Mucke L. Tau reduction prevents Abeta-induced defects in axonal transport. Science. 2010 Oct 8;330(6001):198. PubMed.
- Roberson ED, Halabisky B, Yoo JW, Yao J, Chin J, Yan F, Wu T, Hamto P, Devidze N, Yu GQ, Palop JJ, Noebels JL, Mucke L. Amyloid-β/Fyn-induced synaptic, network, and cognitive impairments depend on tau levels in multiple mouse models of Alzheimer's disease. J Neurosci. 2011 Jan 12;31(2):700-11. PubMed.
- Shipton OA, Leitz JR, Dworzak J, Acton CE, Tunbridge EM, Denk F, Dawson HN, Vitek MP, Wade-Martins R, Paulsen O, Vargas-Caballero M. Tau protein is required for amyloid {beta}-induced impairment of hippocampal long-term potentiation. J Neurosci. 2011 Feb 2;31(5):1688-92. PubMed.
- Ittner LM, Ke YD, Delerue F, Bi M, Gladbach A, van Eersel J, Wölfing H, Chieng BC, Christie MJ, Napier IA, Eckert A, Staufenbiel M, Hardeman E, Götz J. Dendritic function of tau mediates amyloid-beta toxicity in Alzheimer's disease mouse models. Cell. 2010 Aug 6;142(3):387-97. Epub 2010 Jul 22 PubMed.
- Ittner LM, Götz J. Amyloid-β and tau--a toxic pas de deux in Alzheimer's disease. Nat Rev Neurosci. 2011 Feb;12(2):65-72. PubMed.
- Andrews-Zwilling Y, Bien-Ly N, Xu Q, Li G, Bernardo A, Yoon SY, Zwilling D, Yan TX, Chen L, Huang Y. Apolipoprotein E4 causes age- and Tau-dependent impairment of GABAergic interneurons, leading to learning and memory deficits in mice. J Neurosci. 2010 Oct 13;30(41):13707-17. PubMed.
- Dawson HN, Ferreira A, Eyster MV, Ghoshal N, Binder LI, Vitek MP. Inhibition of neuronal maturation in primary hippocampal neurons from tau deficient mice. J Cell Sci. 2001 Mar;114(Pt 6):1179-87. PubMed.
- Takei Y, Teng J, Harada A, Hirokawa N. Defects in axonal elongation and neuronal migration in mice with disrupted tau and map1b genes. J Cell Biol. 2000 Sep 4;150(5):989-1000. PubMed.
- Dawson HN, Cantillana V, Jansen M, Wang H, Vitek MP, Wilcock DM, Lynch JR, Laskowitz DT. Loss of tau elicits axonal degeneration in a mouse model of Alzheimer's disease. Neuroscience. 2010 Aug 11;169(1):516-31. Epub 2010 Apr 29 PubMed.
- Harada A, Oguchi K, Okabe S, Kuno J, Terada S, Ohshima T, Sato-Yoshitake R, Takei Y, Noda T, Hirokawa N. Altered microtubule organization in small-calibre axons of mice lacking tau protein. Nature. 1994 Jun 9;369(6480):488-91. PubMed.
- Caricasole A, Copani A, Caraci F, Aronica E, Rozemuller AJ, Caruso A, Storto M, Gaviraghi G, Terstappen GC, Nicoletti F. Induction of Dickkopf-1, a negative modulator of the Wnt pathway, is associated with neuronal degeneration in Alzheimer's brain. J Neurosci. 2004 Jun 30;24(26):6021-7. PubMed.
- Rosi MC, Luccarini I, Grossi C, Fiorentini A, Spillantini MG, Prisco A, Scali C, Gianfriddo M, Caricasole A, Terstappen GC, Casamenti F. Increased Dickkopf-1 expression in transgenic mouse models of neurodegenerative disease. J Neurochem. 2010 Mar;112(6):1539-51. PubMed.
Other Citations
External Citations
Further Reading
Papers
- Rapoport M, Dawson HN, Binder LI, Vitek MP, Ferreira A. Tau is essential to beta -amyloid-induced neurotoxicity. Proc Natl Acad Sci U S A. 2002 Apr 30;99(9):6364-9. PubMed.
- Roberson ED, Scearce-Levie K, Palop JJ, Yan F, Cheng IH, Wu T, Gerstein H, Yu GQ, Mucke L. Reducing endogenous tau ameliorates amyloid beta-induced deficits in an Alzheimer's disease mouse model. Science. 2007 May 4;316(5825):750-4. PubMed.
- Roberson ED, Halabisky B, Yoo JW, Yao J, Chin J, Yan F, Wu T, Hamto P, Devidze N, Yu GQ, Palop JJ, Noebels JL, Mucke L. Amyloid-β/Fyn-induced synaptic, network, and cognitive impairments depend on tau levels in multiple mouse models of Alzheimer's disease. J Neurosci. 2011 Jan 12;31(2):700-11. PubMed.
- Shipton OA, Leitz JR, Dworzak J, Acton CE, Tunbridge EM, Denk F, Dawson HN, Vitek MP, Wade-Martins R, Paulsen O, Vargas-Caballero M. Tau protein is required for amyloid {beta}-induced impairment of hippocampal long-term potentiation. J Neurosci. 2011 Feb 2;31(5):1688-92. PubMed.
- Ittner LM, Ke YD, Delerue F, Bi M, Gladbach A, van Eersel J, Wölfing H, Chieng BC, Christie MJ, Napier IA, Eckert A, Staufenbiel M, Hardeman E, Götz J. Dendritic function of tau mediates amyloid-beta toxicity in Alzheimer's disease mouse models. Cell. 2010 Aug 6;142(3):387-97. Epub 2010 Jul 22 PubMed.
- Andrews-Zwilling Y, Bien-Ly N, Xu Q, Li G, Bernardo A, Yoon SY, Zwilling D, Yan TX, Chen L, Huang Y. Apolipoprotein E4 causes age- and Tau-dependent impairment of GABAergic interneurons, leading to learning and memory deficits in mice. J Neurosci. 2010 Oct 13;30(41):13707-17. PubMed.
- Dawson HN, Cantillana V, Jansen M, Wang H, Vitek MP, Wilcock DM, Lynch JR, Laskowitz DT. Loss of tau elicits axonal degeneration in a mouse model of Alzheimer's disease. Neuroscience. 2010 Aug 11;169(1):516-31. Epub 2010 Apr 29 PubMed.
- Vossel KA, Zhang K, Brodbeck J, Daub AC, Sharma P, Finkbeiner S, Cui B, Mucke L. Tau reduction prevents Abeta-induced defects in axonal transport. Science. 2010 Oct 8;330(6001):198. PubMed.
News
- APP Mice: Losing Tau Solves Their Memory Problems
- The Plot Thickens: The Complicated Relationship of Tau and Aβ
- San Diego: Pilin’ on the Pyro, Aβ Going Rogue
- Tau’s Synaptic Hats: Regulating Activity, Disrupting Communication
- Honolulu: The Missing Link? Tau Mediates Aβ Toxicity at Synapse
- San Francisco: Gladstone Institute Hosts Tau Powwow
- San Francisco: Making Tau Toxic—Post-translational Changes Galore
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.