Profilin-1 Links Cytoskeleton and RNA Aggregation in ALS
Quick Links
Several proteins linked to amyotrophic lateral sclerosis and frontotemporal dementia show up in stress granules. These natural RNA-protein accumulations protect essential RNAs in times of cellular strife, but cause trouble if they fail to dissipate when the good times return. Stress granules have become a major theme in the study of ALS and related disorders, Robert Brown of the University of Massachusetts Medical School in Worcester noted when he gave the opening address at “RNA Metabolism in Neurological Disease,” a symposium held November 7–8 in San Diego. Researchers have observed disease-associated mutants of TDP-43, FUS, hnRNPA1, hnRNPA2, and ataxin-2 all mired in pathological stress granules. Unexpectedly, Aaron Gitler and Matthew Figley of Stanford University in Palo Alto, California, added another ALS gene to the list: profilin-1, an actin-binding protein. Profilin-1 associates with stress granules in yeast and in mammal cell lines, they reported.
Gitler said he has come to think of stress granules as “crucibles of ALS pathogenesis” because their failure to disassemble may promote neurodegeneration (see Li et al., 2013 and reviewed in Wolozin, 2012). He and Figley offered their results in an oral presentation and poster, respectively.
They did not expect to find themselves studying stress granules when they followed up on the identification of profilin-1 as an ALS gene. ALS-linked profilin-1 mutations occur in the protein’s actin-binding domain, leading researchers to suspect that defects in actin filament assembly or axon outgrowth predispose to the disease, but the mechanism remains a mystery. To get a handle on it, Figley, who works in Gitler’s lab, studied the yeast version of profilin-1, PFY1, which also interacts with actin.
Yeasts that lack PFY1 or express a mutated variant grow at 30 degrees Celsius, but, unlike wild-type yeast, they are inviable at 37 degrees. Adding the human profilin-1 gene allowed the cells to tolerate the higher temperature, reported Figley. He reasoned that he could use the yeast system to assess the functionality of various human profilin-1 variants. The cysteine-71-glycine and methionine-114-threonine substitutions, for example, failed to rescue the deletion strain, whereas the glutamate-117-glycine mutation did, indicating that the first two mutations are fully inactive while E117G might retain some function. This lines up neatly with human results. People with the E117G substitution have no sign of disease, leading scientists to suspect it creates a less severe defect than the C71G and M114T mutations, which turned up only in ALS cases in the original study (see Jul 2012 news story).
To understand profilin’s natural role in yeast, Figley created a set of strains with mutations in PFY1 plus one other gene—for a total of 4,800 crosses—and looked for double mutants that failed to grow. The screen identified three categories of genes that interact with PFY1. One comprised those involved in actin and actin-related cellular activities—no great surprise there. Another consisted of genes related to dynactin, which helps drive intracellular transport. This was interesting, Gitler said, because profilin-1 had no prior link to dynactin. The multipart dynactin complex helps dynein tote cellular cargo along microtubules, and mutations in dynactin components have been linked to motor neuron disease (see Münch et al., 2004; Münch et al., 2005). What was surprising to Gitler and Figley was the third, noncytoskeleton category of genes. About 10 percent of the hits related to stress granules or P-bodies, which are similar mRNA aggregates. Might profilin-1 be important for stress granule formation? To test this hypothesis, Figley stressed various cell types and looked for profilin-1 in the RNA foci.
The researchers treated HeLa cervical cancer cultures with arsenite or dithiothreitol, and observed that profilin-1 co-localized with ataxin-2 and Hu protein R, another stress granule marker. Treatment with cyclohexamide, which prevents RNA from forming into stress granules, prevented the profilin-1 foci (see image below). The same pattern occurred in the human osteosarcoma line U2OS, and in primary mouse cortical neurons. A heat shock also redistributed profilin-1 to RNA foci in the primary neural line. Gitler and Figley concluded that the profilin-1 foci were likely to be stress granules.
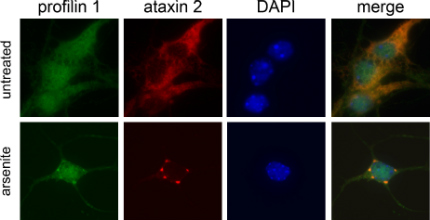
Profiling Stress
In primary cortical mouse neurons under duress, profilin-1 (green) co-localized with the known stress granule marker ataxin 2 (red).
The researchers still need to work out how profilin-1 mutations affect stress granules in ALS. The researchers are collaborating with Paul Taylor at St. Jude Children’s Research Hospital in Memphis, Tennessee, to video granule assembly and disassembly in living cells and sort out how profilin-1 might contribute to motor neuron degeneration. Thus far, ALS-linked profilin-1 mutants appear to accumulate in abnormal RNA aggregates, Gitler said.
Ben Wolozin of Boston University said that in hindsight, he did not find it so surprising that profilin-1 might participate in stress-granule formation. When those structures form, they must rely on the cytoskeleton to deliver components; similarly, the cytoskeleton must pull the granules apart when the RNAs are needed again, he reasoned. He speculated that profilin-1 loss-of-function mutants might be unable to disassemble stress granules, causing long-lasting, pathological granules to accumulate.
Overall, the idea of pathological stress granules has become widely accepted, Wolozin said. Treatments to dissolve pathological stress granules could have therapeutic potential, he added.—Amber Dance.
References
News Citations
Paper Citations
- Li YR, King OD, Shorter J, Gitler AD. Stress granules as crucibles of ALS pathogenesis. J Cell Biol. 2013 Apr 29;201(3):361-72. PubMed.
- Wolozin B. Regulated protein aggregation: stress granules and neurodegeneration. Mol Neurodegener. 2012;7:56. PubMed.
- Münch C, Sedlmeier R, Meyer T, Homberg V, Sperfeld AD, Kurt A, Prudlo J, Peraus G, Hanemann CO, Stumm G, Ludolph AC. Point mutations of the p150 subunit of dynactin (DCTN1) gene in ALS. Neurology. 2004 Aug 24;63(4):724-6. PubMed.
- Münch C, Rosenbohm A, Sperfeld AD, Uttner I, Reske S, Krause BJ, Sedlmeier R, Meyer T, Hanemann CO, Stumm G, Ludolph AC. Heterozygous R1101K mutation of the DCTN1 gene in a family with ALS and FTD. Ann Neurol. 2005 Nov;58(5):777-80. PubMed.
Further Reading
Papers
- Ramaswami M, Taylor JP, Parker R. Altered ribostasis: RNA-protein granules in degenerative disorders. Cell. 2013 Aug 15;154(4):727-36. PubMed.
- Buchan JR, Kolaitis RM, Taylor JP, Parker R. Eukaryotic stress granules are cleared by autophagy and Cdc48/VCP function. Cell. 2013 Jun 20;153(7):1461-74. PubMed.
- Kim HJ, Kim NC, Wang YD, Scarborough EA, Moore J, Diaz Z, Maclea KS, Freibaum B, Li S, Molliex A, Kanagaraj AP, Carter R, Boylan KB, Wojtas AM, Rademakers R, Pinkus JL, Greenberg SA, Trojanowski JQ, Traynor BJ, Smith BN, Topp S, Gkazi AS, Miller J, Shaw CE, Kottlors M, Kirschner J, Pestronk A, Li YR, Ford AF, Gitler AD, Benatar M, King OD, Kimonis VE, Ross ED, Weihl CC, Shorter J, Taylor JP. Mutations in prion-like domains in hnRNPA2B1 and hnRNPA1 cause multisystem proteinopathy and ALS. Nature. 2013 Mar 28;495(7442):467-73. PubMed.
- Bosco DA, Lemay N, Ko HK, Zhou H, Burke C, Kwiatkowski TJ Jr, Sapp P, McKenna-Yasek D, Brown RH Jr, Hayward LJ. Mutant FUS proteins that cause amyotrophic lateral sclerosis incorporate into stress granules. Hum Mol Genet. 2010 Nov 1;19(21):4160-75. Epub 2010 Aug 10 PubMed.
Annotate
To make an annotation you must Login or Register.
Comments
No Available Comments
Make a Comment
To make a comment you must login or register.